Организм как открытая саморегулирующаяся система. Единство организма и внешней среды. Гомеостаз
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
Возбуждение по нервному волокну распространяется в обе стороны от места его возникновения, то есть как центробежно, так и центростремительно. Нервные волокна обладают лабильностью (пропускной способностью) Лабильность – это способность воспроизводить определённое количество потенциалов действия в единицу времени в соответствии с ритмом действующего раздражителя. Мерой лабильности является максимальное количество ПД, которое способна воспроизвести структура без искажения ритма. Лабильность определяется длительностью пика потенциала действия, в частности, продолжительностью фазы абсолютной рефрактерности. Поскольку абсолютная рефрактерность у нервного волокна самая короткая, то нервное волокно способно воспроизвести от 500 до 1000 импульсов в секунду. Лабильность скелетной мышцы 200-300 имп./с 43. Система опоры и движения, её исполнительные органы, роль в организме. Общим свойством всего живого и основой активного поведения является движение. Органом движения является мышечный аппарат, который включает 3 вида мышц: скелетные, гладкие и сердечную мышцы. Мышцы выполняют следующие функции: 1. Создание позы и удержание тела в пространстве, преодоление инерции. 2. Двигательная функция внутренних органов (моторная функция кишечника, сократительная функция сердца, обеспечение дыхания за счёт сокращения дыхательных мышц). 3. Эффекторный механизм мыслительной (произносимая речь) и поведенческой деятельности. 4. Преобразование химической энергии макроэргических соединений в механическую, тепловую, электрическую энергию.
44. Биоэлектрические явления в мышечном волокне. Особенности потенциала покоя и потенциала действия. Физиологические свойства скелетных мышц: Общие
Частные
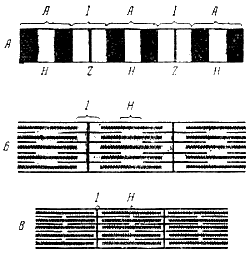
Одиночное сокращение – возникает при действии одиночным раздражителем (прямое раздражение), или через иннервирующий её двигательный нерв (непрямое раздражение). Тетаническое (суммированное) сокращение – длительное и сильное сокращение мышцы в ответ на ритмическое раздражение. Электромеханическое сопряжение это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску цикла поперечных мостиков (последовательность событий, начиная от связывания поперечного мостика с тонким филаментом и до момента, когда система готова к повторению процесса). • происходит посредством распространения потенциала действия по мембранам поперечной системы в глубь клетки. • При этом возбуждение быстро проникает вглубь волокна, переходит на продольную систему и, в конечном счете, вызывает высвобождение кальция из терминальных цистерн во внутриклеточную жидкость, окружающую миофибриллы, что и ведет к сокращению Плазматическая мембрана скелетных мышц электрически возбудима и способна генерировать распространяющийся потенциал действия посредством механизма, аналогичного тому, который действует в нервных клетках. Потенциал действия в волокне скелетной мышцы длится 1-2 мс и заканчивается раньше, чем появятся какие-либо признаки механической активности. Начавшаяся механическая активность может продолжаться более 100 мс. Электрическая активность плазматической мембраны не оказывает прямого влияния на сократительные белки, а вызывает повышение цитоплазматической концентрации ионов Са2+, которые продолжают активировать сократительный аппарат и после прекращения электрического процесса. Величина мембранного потенциала гладкомышечного волокна зависит от состояния мышцы в данный момент. В норме в состоянии покоя внутриклеточный потенциал обычно составляет около -50-60 мВ, т.е. он примерно на 30 мВ менее отрицателен, чем в скелетной мышце. Мембранный потенциал покоя мышечного волокна составляет 80-90 мв, а потенциал действия - 120-135 мв. Условием возникновения потенциала действия является критическая деполяризация мембраны. При непрямом раздражении мышцы критическая деполяризация мембраны осуществляется ацетилхолином, выделяющимся в нервных окончаниях при поступлении к ним нервного импульса. Возникший в области нервно-мышечного соединения потенциал действия распространяется далее вдоль всего мышечного волокна. При прямом раздражении электрическим током потенциал действия в мышечном волокне возникает под катодом и распространяется из этой точки по волокну. Длительность потенциала действия мышечного волокна - 3-5 мсек, т.е. 5-10 раз больше продолжительности потенциала действия двигательных нервных волокон, иннервирующих мышцу. Поэтому максимальное число импульсов, которое может воспроизводить мышца, не превышает 200-250 в секунду. В возникновении потенциала покоя в скелетном мышечном волокне главную роль играют ионы Кֺ, диффундирующие из клетки в окружаю среду, и ионы Сl' диффундирующие в противоположном направлении. Потенциал действия обусловлен повышением натриевой проницаемости мембраны и потоком ионов Naֹ внутрь волокна. 45. Изменения возбудимости скелетной мышцы во время одиночного цикла возбуждения. • Возбудимость мышц в ходе одиночного сокращения меняется. • Во время развития ПД, т.е. в начале сокращения мышцы наступает абсолютная рефрактерная фаза (состояние невозбудимости, длительностью 3-5 мсек.). • За ней следует фаза относительной рефрактерности, а затем • возникает фаза повышенной возбудимости (примерно 20 мсек.). • Длительность этих периодов различна у животных разных видов и зависит от функционального состояния нервно-мышечного аппарата. 46. Ультрамикроскопическая структура миофибрилл в покое и при сокращении. Понятие о саркомере. Ультрамикроскопическая структура миофибрилл в покое и при сокращении. Миофибриллы скелетных мышц оптически неоднородны: состоят из изотропных дисков I и анизотропных дисков А. Причина этой неоднородности была раскрыта в работах А. X и Г. Хаксли, которые применили для исследования структуры мышечного волокна электронную микроскопию, рентгеноструктурный анализ, фазовоконттрастную и интерференционную микроскопию в сочетании с гистохимическими методами. Было установлено, что каждая из миофибрилл мышечного волокна диаметром около 1 мк состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимеризованные молекулы белков миозина и актина. Миозиновые протофибриллы, или, как их обычно принято обозначать, нити, вдвое толще актиновых. Их диаметр составляет примерно 100 Å. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 147, Б). Благодаря этому диски I, как показывет рисунок, состоят только из актиновых нитей, а диски А из миозиновых и актиновых. Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. Мембрана Z, проходя через середину диска I, скрепляет между собой эти нити. Важным компонентом ультрамикроскопической структуры миофибрилл являются также многочисленные поперечные мостики, соединиющие между собой миозиновые нити с актиновыми. При сокращении мышечного волокна, согласно теории А. Хаксли и Г. Хаксли, указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего длина дисков I укорачивается, а диски А сохраняют свой размер. Почти исчезает лишь светлая полоска Н, так Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая зависимость. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем постепенно падает. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю.
Саркомер (повторяющийся сегмент миофибриллы) состоит из двух половин светлого, оптически изотропного диска (I) и одного темного, анизотропного (А) диска (см. рис. 1.51). Электронно—микроскопический и биохимический анализы показывают, что у позвоночных темный диск сформирован параллельным пучком толстых (диаметром порядка 10 нм) миозиновых нитей, имеющих длину около 1,6 мкм. Молекулярная масса белка миозина составляет 500 килодальтон (кДа). На нитях миозина расположены выступы — головки миозиновых молекул длиной 20 нм. В светлых дисках имеются тонкие нити (диаметром 5 нм, длиной 1 мкм), построенные из белка актина (молекулярная масса 42 кДа), а также тропомиозина и тропонина. В районе Z—линии, разграничивающей соседние саркомеры, пучок тонких нитей скреплен Z—мембраной. Соотношение тонких и толстых нитей в саркомере составляет 2:1. Миозиновые и актиновые нити саркомера расположены так, что тонкие нити могут свободно входить между толстыми, т. е. «задвигаться» в А— диск, что и происходит при сокращении мышцы. В силу этого длина светлой части саркомера —I—диска — может быть разной: при пассивном растяжении мышцы она увеличивается до максимума, при сокращении может уменьшаться до нуля. 47. Механизм мышечного сокращения и расслабления. Теория «скольжения» нитей. Химические и тепловые изменения в мышечном волокне. Скелетная мускулатура иннервируется импульсом, приходящим по СНС. Это волокно выходит из передних рогов СМ. Одно волокно может иннервировать несколько мышечных клеток. Одна клетка иннервируется одним волокном. Выделяется медиатор ацетилхолин и действует на н-холинорецепторы. Возникший ПД деполяризует мембрану СПР, в результате чего открываются потенциалуправляемые Ca-каналы. Ca попадает внутрь клетки. После выхода ионов Ca из СПР, [Ca2+] в ЦП увеличивается. Как только Ca2+ связывается с тропонином, белок тропонин меняет геометрию. В результате актиновый центр освобождается от тропонина, туда присоединяются миозиновые головки. Происходит расщепление АТФ на АДФ+Энергия. Энергия тратится на поворот головки миозина на90 градусов вместе с актиновым центром. После этого перемещения головка миозина отсоединяется от актинового центра и эластическими центрами возвращается исходное положение. После чего снова приходит вместе с молекулой АТФ. В основе мышечного сокращения лежит теория скользящих нитей. Основное положение теории скользящих нитей – во время скольжения (сокращения) сами актиновые и миозиновые нити не укорачиваются, так как ширина диска А остается при сокращении постоянной, а І-диски и Н-зоны становятся более узкими или совсем исчезают. Длина протофибрилл не изменяется и при растяжении мышцы. Вместо этого пучки тонких нитей, скользя, выходят из промежутков между толстыми нитями, так что степень их перекрытия уменьшается. Мышечное расслабление начинается с того, что закрываются Ca-каналы СПР и начинают работать каналы-насосы Ca на СПР. На перенос Ca в СПР тратится АТФ. В результате этого [Ca2+] в ЦП резко снижается и белок тропонин теряет Ca, принимает исходную геометрию и закрывает актиновый центр. Начинается процесс расслабления. Потенциалы действия, возникающие на мембране мышечного волокна, приводят к ряду химических процессов, которые завершаются механической реакцией волокна. Связь между электрическими процессами, происходящими на мембране, и механической реакцией миофибрилл обеспечивается ионами кальция (Са++). В состоянии покоя они находятся внутри трубочек и полостей, пронизывающих мышечное волокно, а при возбуждении, в связи с повышением проницаемости мембран этих трубочек, выходят из них и оказываются вблизи миофибрилл. Ионы Са++ влияют на белок миозин, который является не только сократительным белком, но и обладает свойствами фермента. Когда волокна находятся в состоянии покоя, миозин как фермент неактивен. При воздействии же ионов Са++ его ферментативные свойства активизируются, и он начинает расщеплять богатое энергией вещество - аденозинтрифосфорную кислоту (АТФ). Эа счет освобождающейся химической энергии происходит сокращение миофибрилл, т. е. втягивание актиновых протофибрилл в промежутки между миозиновыми. Расслабление мышечного волокна связано с удалением Са++ от миофибрилл. АТФ является непосредственным источником энергии для мышечного сокращения. Однако запас этого вещества весьма ограничен. Возможность совершать работу в течение более или менее длительного времени связана с процессами непрерывного восстановления количества АТФ в мышце. Таким образом, расщепление АТФ это лишь первое звено в цепи химических реакций, которые происходят в мышце при ее деятельности. Вслед за ним совершаются реакции, обеспечивающие ресинтез АТФ. Ресинтез АТФ заключается в присоединении к аденозиндифосфорной кислоте (АДФ), образовавшейся при распаде АТФ, молекулы фосфорной кислоты. Эта реакция требует энергии. В мышце имеются содержащие энергию вещества. (Есть они и в других органах, откуда могут доставляться мышце кровью.) К ним относятся углеводы, жиры, белки. Но заключенная в них энергия освобождается только при их расщеплении, которое происходит под влиянием ферментов. Ресинтез АТФ совершается двояким путем: за счет расщепления веществ без участия кислорода (анаэробные процессы) и с участием кислорода (аэробные процессы). Ресинтез АТФ анаэробным путем происходит в первую очередь за счет креатинфосфорной кислоты (КрФ), которая, реагируя с АДФ, отдает ей фосфорную кислоту, обеспечивая восстановление АТФ, Однако запас КРФ в мышце тоже ограничен. Несколько позднее включаются анаэробные реакции расщепления углеводов - гликогена, глюкозы, которых в организме значительно больше, чем Крф. При этом в организме накапливаются промежуточные продукты распада углеводов (молочная кислота и др.), что снижает его работоспособность. Ресинтез АТФ аэробным путем происходит за счет окислительного распада углеводов, жиров и других веществ до углекислоты и воды. При этом освобождается большое количество энергии, обеспечивающей весьма эффективный процесс ресинтеза АТФ. В связи с этим при мышечной деятельности возникает потребность в повышенном поглощении кислорода. Итак, в мышце происходят сложные химические процессы, многие из которых имеют обратимый характер: вещества расщепляются и восстанавливаются. АТФ, будучи первичным источником энергии для мышечного сокращения, может при благоприятных условиях полностью восстанавливаться. Восстановлению подвергаются и КРФ, и частично углеводы. В конечном итоге расходуется то количество органических веществ, которое распадается до углекислоты и воды. Химические реакции, происходящие в мышце при возбуждении, непосредственно связаны с энергетическими процессами. Распад АТФ приводит к превращению потенциальной химической энергии в механическую; ресинтез АТФ и других веществ связан с переносом энергии с одних соединений на другие. Энергетические процессы всегда сопровождаются образованием тепла. В мышце химическая энергия превращается в механическую непосредственно. Следовательно, мышца является не тепловым двигателем, а химическим. Тем не менее и в ней образуется тепло. Часть его представляет собой неизбежную утечку энергии при превращении химической энергии в механическую, а также при ресинтезе веществ; часть тепла образуется в связи с сокращением мышцы (в результате внутреннего трения перемещающихся частей мышечных волокон), часть - при ее расслаблении (в результате вторичного процесса превращения энергии - перехода механической энергии в тепловую). Тепло, образующееся в мышцах, имеет большое значение для поддержания температуры. Таким образом: Последовательность основных процессов при мышечном сокращении:
Последовательность основных процессов при мышечном расслаблении:
48. Одиночное мышечное сокращение и его характеристика. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение. Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока. Фазы одиночного сокращения: - латентная (10 мс) - укорочения (50 мс) - расслабления (50 мс) 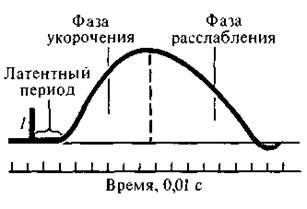 49. Сопоставить фазы потенциала действия с фазами изменения возбудимости и одиночного цикла сокращения. Отметить особенности рефрактерного периода. При нанесении одиночного порогового и сверхпорогового раздражения на мышцу, мышца сокращается, а затем расслабляется, то есть наблюдается одиночное сокращение мышц. Имеет место ряд последовательных явлений: • после нанесения раздражения сокращение мышцы наступает не сразу, а через некоторый интервал 2,5 мс (латентный период). • Реакция мышцы на раздражение начинается с генерации ПД продолжительностью 3-5 мс и в это же время • начинается сокращение мышцы, приблизительно 50 мсек. (фаза сокращения), • затем следует фаза расслабления, по продолжительности либо равна или может быть несколько больше фазного сокращения. Длительность зависит от морфофункциональных свойств. • Так, у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения равна 7-10 мс, а у медленных волокон камбаловидной мышцы равна 50-100 мс. 50. Суммация сокращений, виды суммации. Условия суммации. В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы. Чаще одиночные сокращения суммируются. Суммация – образование ПД в фазе расслабления. Суммация это сложение 2-х последовательных сокращений при нанесении на нее 2-х пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефрактерного периода. Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда он уже начала расслабляться. Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения.(рис 1,2). Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить. Например нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности. 51. Тетанус, его виды. Теории тетануса. Оптимум и пессимум частоты раздражения. В естественных условиях к мышечным волокнам поступают не одиночные, а ряд нервных импульсов, на которые мышца отвечает длительным, тетаническим сокращением, или тетанусом. К тетаническому сокращению способны только скелетные мышцы. Гладкие мышцы и поперечнополосатая мышца сердца не способны к тетаническому сокращению из-за продолжительного рефрактерного периода. Тетанус возникает вследствие суммации одиночных мышечных сокращений. Чтобы возник тетанус, необходимо действие повторных раздражений (или нервных импульсов) на мышцу еще до того, как закончится ее одиночное сокращение. Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться, то возникает зубчатый тип сокращения (зубчатый тетанус). Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, то есть происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого тетануса. Гладкий тетанус – нормальное рабочее состояние скелетных мышц обусловливается поступлением из ЦНС нервных импульсов с частотой 40-50 в 1с. Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с. Если мышца получает 10-20 нервных импульсов в 1с, то она находится в состоянии мышечного тонуса, т.е. умеренной степени напряжения. 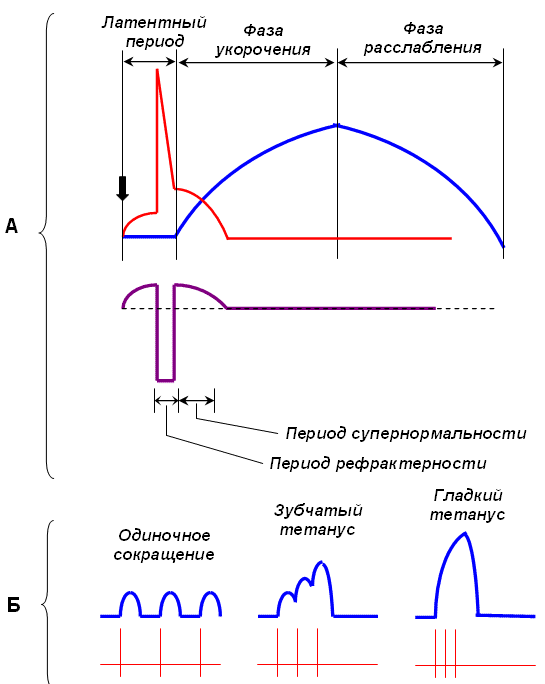 Теории тетануса: • Механическая теория суперпозиции сокращений (Г. Гельмгольц). При этом каждое последующее сокращение формируется как бы с исходного уровня. • Теория изменения состояния мышцы при воздействиях (Н. Е. Введенский), согласно которой предыдущее состояние мышцы определяет ее ответ на последующее раздражение. Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается. 52. Морфо-функциональные особенности гладких мышц. Гладкие мышцы и их особенности
Морфологические особенности:
Физиологические особенности:
Раздражители, вызывающие сокращение гладких мышц:
Особенности электрических процессов гладких мышц
53. Моторные синапсы, строение, функциональные свойства, механизм передачи возбуждения. Фармакологическая коррекция работы мионеврального синапса. |