Организм как открытая саморегулирующаяся система. Единство организма и внешней среды. Гомеостаз
 Скачать 2.85 Mb. Скачать 2.85 Mb.
|

|
Нервно-мышечный синапс - соединение концевой ветви аксона мотонейрона спинного мозга с мышечной клеткой.
Постсинаптические структуры: постсинаптическая мембрана (мышечной клетки), субсинаптическая мембрана (постсинаптической мембраны). По структуре и функции нервно-мышечный синапс является типичным химическим синапсом. Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой. Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинап-тическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с преси-наптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холи-нэстеразой (ХЭ), которые находятся на постсинапти-ческой мембране. Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом: ХР—ХЭ—ХР—ХЭ—ХР—ХЭ. ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины. Затем происходит суммация МПКП. В результате сум-мации образуется ВПСП – возбуждающий постсинап-тический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна. ХЭ + АХ = разрушение АХ до холина и уксусной кислоты. В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинап-тической мембраной лопается, и ее содержимое в виде 1 кванта (10000 молекул) АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН. Мионевральный синапс обладает следующими основными свойствами. 1. Синапс проводит возбуждение только в одном направлении – в направлении от пресинаптической мембраны к постсинаптической. 2. В синапсе имеет место синаптическая задержка возбуждения, т. е. скорость проведения возбуждения по синапсу значительно меньше, чем по нервному волокну. Это связано с определенной продолжительностью времени, необходимого для выделения медиатора и взаимодействия его с рецепторами. 3. В синапсе отмечается облегчение проведения каждого последующего возбуждения, что, по всей вероятности, связано с накоплением медиатора в синаптической щели. 4. При длительном возбуждении синапса в нем может наблюдаться снижение чувствительности рецепторов к медиатору, обусловленное закрытием части натриевых каналов, за счет включения системы инактивации. 5. В синапсах быстро развивается процесс утомления, связанный с быстрым метаболическим истощением запасов медиатора в везикулах пресинаптических утолщений. Фармакологическая коррекция: - миастения Гравис – образование антител к АХ-рецепторам > АХ не связывается с рецепторами. Лечение – АХэстеразные препараты, блокирующие активность ацетилхолинэстеразы > усиление действия АХ, что позволяет ему свзяваються с заблокированными рецепторами - ботулотоксины (Ботокс, LANTOX) – местные релаксанты. Блокируют высвобождение ацетилхолина из окончания нервного волокна в области нервно-мышечного синапса. Предполагается, что ингибирование высвобождения ацетилхолина приводит к прорастанию новых нервных окончаний, вследствие чего восстанавливается нервно-мышечный синапс.:
54. Структурно-функциональные особенности скелетных мышц. Понятие о моторной единице. Виды моторных единиц. Анатомической и функциональной единицей скелетных мышц является моторнаяединица - двигательный нейрон и иннервируемая им группа мышечных волокон. Импульсы, посылаемые мотонейроном, приводят в действие все образующие ее мышечные волокна. По характеру возбуждения, возникающего в мышечных волокнах все нейромоторные единицы делятся на 2 группы. Фазные нейромоторные единицы - образуются альфа-мотонейронами. Это самые крупные мотонейроны (диаметр 10-20 мкм). Скорость проведения возбуждения по отростку этого аксона - 120 м/с. Аксон обеспечивает одиночную иннервацию, образуя на каждом волокне 1-2 синапса. В каждом синапсе при возбуждении возникает достаточный потенциал концевой пластинки, который обеспечивает возникновение потенциала действия в мышце (импульсное возбуждение). Лучше работают под действием одиночного возбуждения (импульса), при котором возникает распространяющееся возбуждение. Среди фазных нейромоторных единиц выделяют быстрые и медленные.
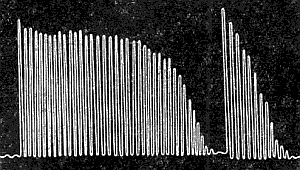
Тонические нейромоторные единицы - образуются гамма-мотонейронами передних рогов спинного мозга (самые мелкие, диаметр 4-6 мкм). Скорость проведения импульса по отросткам этих нейронов - 30 м/с, в синапсах возникает местный потенциал концевой пластинки (местное возбуждение). Аксоны тонических нейронов обеспечивают множественную иннервацию, т. е. на каждом мышечном волокне образуются несколько десятков синапсов и за счет суммации местных потенциалов и возникает потенциал действия. Возбуждение тонической нейромоторной единицы происходит под влиянием серии импульсов (частота около 10 Гц). Тонические нейромоторные единицы обеспечивают медленное сокращение мышц, участвуют в возникновении тонуса. Скелетные мышцы образуются различными мышечными волокнами, которые входят в состав фазных и тонических нейромоторных единиц. Попеременное включение нейромоторных единиц обеспечивает изменение функционального состояния мышцы. В состоянии покоя работают тонические нейромоторные единицы, в состоянии активности - фазные нейромоторные единицы. Кроме двигательной иннервации присутствует вегетативная. Все скелетные мышцы получают импульсы из симпатической нервной системы, которая регулирует обменные процессы. Скелетные мышцы состоят из большого количества мышечных волокон. Волокно поперечнополосатой мышцы имеет вытянутую форму, диаметр его от 10 до 100 мкм, длина волокна от нескольких сантиметров до 10-12 см. Мышечная клетка окружена тонкой мембраной – сарколеммой, содержит саркоплазму (протоплазму) и многочисленные ядра. Сократительной частью мышечного волокна являются длинные мышечные нити – миофибриллы, состоящие в основном из актина, проходящие внутри волокна от одного конца до другого, имеющие поперечную исчерченность. Основной функцией скелетных мышц является сокращение, которое выражено различными движениями человека. Скелетные мышцы выполняют также рецепторную, обменную и терморегулирующую функции. Они образуются большим количеством многоядерных мышечных волокон. Сократительной частью мышечного волокна являются длинные мышечные нити — миофибриллы, которые проходят внутри волокна от одного конца к другому и имеют поперечную очерченность. Последняя образована чередованием темных (анизотропных) А-дисков и светлых (изотропных) 1-дисков (см. рис. 53). Через середину 1-диска проходит Z-линия; две соседние Z-линии ограничивают саркомер, структурно-фунциональную единицу. При электронной микроскопии волокон видно, что в составе А-диска есть более светлый участок (Н-зона), а в центре этот диск пересекает темная полоса — М-линия. Темный диск образован толстыми нитями белка миозина, а светлый 1-диск — тонкими нитями белка актина. В мышечном волокне содержатся также фибриллярный палочковидный белок — тропомиозин и глобулярный белок — тропонин. 55. Режимы сокращений скелетных мышц (изотоническое, изометрическое, ауксотоническое). Лестничная зависимость между силой раздражения и амплитудой сокращения скелетной мышцы. Во время выполнения работы мышца может сокращаться: • изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте; • изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы; • ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе. Амплитуда одиночного сокращения изолированного мышечного волокна от силы раздражения не зависит, т.к. подчиняется закону «все или ничего». А сокращение целой мышцы, состоящей из множества волокон, при ее прямом раздражении находится в большей зависимости от силы раздражения. При пороговой силе тока в реакцию вовлекается лишь небольшое число волокон, поэтому сокращение мышцы едва заметно. С увеличением силы раздражения число волокон, охваченных возбуждением, возрастает, сокращение усиливается до тех пор, пока все волокна не оказываются сокращенными («максимальное сокращение»). После этого усиление раздражающего стимула на амплитуду сокращения мышцы не влияет. Таким образом, скелетная мышца отвечает на раздражение градуально (в зависимости от раздражительного стимула). 56. Сила мышцы. Факторы, влияющие на силу мышцы. Максимальная сила мышц – это величина максимального напряжения, которое может развить мышца. Она зависит от строения мышцы, ее функционального состояния, исходной длины, пола, возраста, степени тренированности человека. Сила сокращения скелетной мышцы определяется 2 факторами: • числом моторных единиц, участвующих в сокращении; • частотой сокращения мышечных волокон. Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения. Для сравнения силы различных мышц определяют их удельную или абсолютную силу. Она равна максимальной, деленной на кв. см. площади поперечного сечения мышцы. Удельная сила икроножной мышцы человека составляет 6,2 кг/см2, трехглавой – 16,8 кг/см2, жевательных – 10 кг/см 2. 57. Работа мышцы при разных нагрузках. Правило средних нагрузок. Работа мышцы – её способность сокращаться. Виды работы скелетной мышцы: • динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве; • статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении; • динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела. Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках. 58. Утомление мышц, теории утомления. Утомление изолированной мышцы. Утомление нервно-мышечного препарата. Утомление моторной единицы в условиях организма. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов. В прошлом веке, на основании опытов с изолированными мышцами, было предложено 3 теории мышечного утомления. 1.Теория Шиффа: утомление является следствием истощения энергетических запасов в мышце. 2.Теория Пфлюгера: утомление обусловлено накоплением в мышце продуктов обмена. 3.Теория Ферворна: утомление объясняется недостатком кислорода в мышце. Утомление изолированной мышцы можно вызвать ее ритмическим раздражением. В результате этого сила сокращений прогрессирующе уменьшается (рис) . Чем выше частота, сила раздражения, величина нагрузки тем быстрее развивается утомление. При утомлении значительно изменяется кривая одиночного сокращения. Увеличивается продолжительность латентного периода, периода укорочения и особенно периода расслабления, но снижается амплитуда (рис.) . Чем сильнее утомление мышцы, тем больше продолжительность этих периодов. В некоторых случаях полного расслабления не наступает. Развивается контрактура. Это состояние длительного непроизвольного сокращения мышцы. Работа и утомление мышц исследуются с помощью эргографии. В условиях организма развитие утомления самой мышцы - явление крайне редкое, так как прежде всего утомление развивается в структурах центральной нервной системы и нервно-мышечном синапсе. Однако независимо от того, где произошло утомление, зарегистрированная кривая мышечного утомления носит одинаковый характер. Так, на свежем нервно-мышечном препарате при длительном ритмическом раздражении нерва даже в редком ритме можно получить характерную кривую утомления мышцы. Некоторое время после начала раздражения амплитуда сокращения мышцы увеличивается. Наблюдаемое явление называется "лестницей Боудича" и объясняется активированием процессов обмена в результате перехода мышцы в рабочее состояние. После стадии лестницы амплитуда мышечных сокращений держится некоторое время на постоянном уровне - стадия плато. Эта стадия сменяется стадиями быстрого и медленного спада, во время которых амплитуда сокращений уменьшается, так как наступает утомление. Одновременно с развитием утомления в мышце возникает контрактура - явление, связанное с замедлением процессов расслабления. Однако главная роль в утомлении двигательного аппарата принадлежит моторным центрам ЦНС. В прошлом веке И.М.Сеченов установил, что если наступает утомление мышц одной руки, то их работоспособность восстанавливается быстрее при работе другой рукой или ногами. Он считал, что это связано с переключением процессов возбуждения с одних двигательных центров на другие. Отдых с включением других мышечных групп он назвал активным. В настоящее время установлено, что двигательное утомление связано с торможением соответствующих нервных центров, в результате метаболических процессов в нейронах, ухудшением синтеза нейромедиаторов, и угнетением синаптической передачи. 59. Кривая утомления. Эргография. Локальное и общее утомление. Пассивный отдых. Роль активного отдыха по Сеченову. 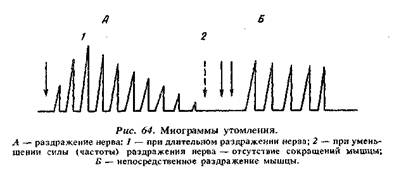 Эргография. Для изучения мышечного утомления у человека в лабораторных условиях пользуются эргографами — приборами для записи амплитуды движения, ритмически выполняемого группой мышц. Примером такого прибора может служить эргограф Моссо, записывающий движение нагруженного пальца при сгибании и разгибании и дающий суммарные сведения о работе собственного сгибателя этого пальца и общего сгибателя всех пальцев руки. Исследуемый, сгибая и разгибая палец, поднимает и опускает подвешенный к пальцу груз в ритме ударов метронома. Особый интерес представляют эргографы, воспроизводящие те или иные рабочие движения человека. Первым таким прибором был эргограф, примененный И. М. Сеченовым для изучения рабочих движений при пилке ручной пилой. Меняя величину груза и частоту ударов метронома, можно установить тот ритм и груз, при которых данный индивидуум в данных условиях эксперимента выполняет наибольшую работу в кратчайший срок.
Утомление, возникающее при физической работе, в которую вовлечены обширные мышечные группы, называется общим. Для общего утомления характерно нарушение регуляторной функции ЦНС, координации двигательной и вегетативной функций, снижение эффективности волевого контроля за качеством выполнения движений. Общее утомление сопровождается расстройствами вегетативных функций: неадекватным нагрузке увеличением ЧСС, падением пульсового давления, уменьшением легочной вентиляции. Субъективно это ощущается как резкий упадок сил, одышка, сердцебиение, невозможность продолжать работу. Когда чрезмерная нагрузка падает на отдельные мышечные группы, развивается так называемое локальное утомление. В отличие от общего утомления, при локальном утомлении страдает не столько центральный аппарат управления, сколько местные структурные элементы регуляции движений: терминали двигательных нервов, нервно-мышечный синапс. Нарушения в нервно-мышечной передаче возбуждения развиваются задолго до того, как сами исполнительные приборы перестают нормально функционировать. В пресинаптической мембране уменьшается количество ацетилхолина, вследствие чего падает потенциал действия постсинаптической мембраны. Происходит частичное блокирование эфферентного нервного сигнала, передаваемого на мышцу. Сократительная функция мышцы ухудшается. Различают два вида отдыха: пассивный и активный. Пассивный — это полный покой. Активный отдых представляет собой отдых от деятельности, вызвавшей утомление. Долгое время господствовало мнение, что быстро восстановить работоспособность можно только при полном покое организма. И. М. Сеченов, исследуя рабочие движения человека, опроверг это представление. Он сравнил быстроту восстановления работоспособности утомленной правой руки при полном покое и в случае, когда правая рука отдыхала, а левая работала. Оказалось, что во втором случае работоспособность восстанавливается быстрей. Происходит это потому, что обратный поток импульсов, идущий от левой руки в центральную нервную систему, возбуждает утомленные (заторможенные) нервные клетки, управляющие работой уставшей руки. И восстановление происходит быстрее. Значит, под влиянием нервных импульсов от неутомленных органов работоспособность утомленных участков мозга восстанавливается быстрее. При полном покое в центральную нервную систему импульсы, возбуждающие заторможенные нервные клетки, не поступают. Вот почему активный отдых более эффективен, чем полный покой. 60. Адаптационно-трофический феномен Орбели-Гинецинского. Роль высших отделов ЦНС в развитии утомления. Если стимуляцией двигательного нерва довести скелетную мышцу до утомления, а затем одновременно раздражать симпатический нерв, то работоспособность утомленной мышцы повышается (феномен Орбели-Гинецинского) - адаптационно-трофическое действие симпатической нервной системы). Мышечная работа - это целостная деятельность всего организма. Функциони рование организма как целого и его взаимодействие с внешним миром осуществляется посредством нервной системы при веду щей роли ее высшего отдела — коры больших полушарий. Утомление организма вследствие мышечной работы является прежде всего результатом сдвигов в функциональном состоянии центральной нервной системы. Исследования отечественных физиологов — И. М. Сеченова, И. П. Павлова, Н. Е. Введенского, А. А. Ухтом ского, Л. А. Орбели, Г. В. Фольборта и др. — убедительно обосно вывают то важное положение, что в возникновении и развитии утомления нервная система играет ведущую роль. Утомление организма при мышечной работе, прежде всего, связано с утомлением центральной нервной системы, так как интенсивная мышечная деятельность является в то же время и интенсивной деятельностью нервных центров. Последняя в результате длительной напряженной работы нарушается. Выражением этого нарушения является изменение нормального взаимоотноше ния процессов возбуждения и торможения, причем тормозной процесс начинает преобладать. В результате расстраивается нор мальное течение рефлекторных процессов, нарушаются регуляция вегетативных функций и координация движений, двигательный аппарат постепенно приходит в недеятельное состояние. Нервная система наиболее чувствительна к изменениям вну тренней среды. Такие факторы утомления, как накопление в крови продуктов работы клеток, уменьшение содержания в крови сахара, недостаток при некоторых условиях кислорода в крови, понижают работоспособность организма не прямо, а глав ным образом опосредствованно — через центральную нервную систему. Эти возможности коры больших полушарий и других отделов мозга, осуществляемые через посредство интрацентральных пу тей и вегетативных нервов, реализуются с помощью регулирую щих влияний на все органы и ткани, в том числе также и на центральную нервную систему. В активизации этих влияний ведущая роль принадлежит рефлекторным реакциям, возникающим при действии самых разнообразных сигнальных раздражителей. 61. Методы изучения функций центральной нервной системы. Электроэнцефалография. Существуют следующие методы исследования функций ЦНС: 1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом. 2. Метод экстирпации (удаления) или разрушения участков мозга. 3. Метод раздражения различных отделов и центров мозга. 4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием. 5. Электрофизиологические методы: а. электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г. Бергером. б. регистрация биопотенциалов различных нервных центров; используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро. в. метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков; 6. метод внутримозгового введения веществ с помощью микроинофореза; 7. хронорефлексометрия – определение времени рефлексов. Электроэнцефалография (ЭЭГ)-это регистрация электрической активности мозга с поверхности кожи головы. Впервые ЭЭГ человека зарегистрировал в 1929 г. немецкий психиатр Г.Бергер. При снятии ЭЭГ на кожу накладывают электроды, сигналы от которых усиливаются и подаются на осциллограф и пишущее устройство. В норме регистрируются следующие типы спонтанных колебаний: 1. Альфа-ритм. Это волны с частотой 8-13 Гц. Наблюдается в состоянии бодрствования, полного покоя и при закрытых глазах. Если человек открывает глаза a-ритм сменяется b-ритмом. Это явление называется блокадой a-ритма. 2. Бета-ритм. Его частота от 14 до 30 Гц. Наблюдается при деятельном состоянии мозга и учащается по мере повышения интенсивности умственной работы. 3. Тета-ритм (θ-ритм). Колебания с частотой 4-8 Гц. Регистрируется во время засыпания, поверхностного сна и неглубоком наркозе. 4. Дельта-ритм. Частота 0,5-3,5 Гц. Наблюдается при глубоком сне и наркозе. Чем ниже частота ритмов ЭЭГ, тем больше их амплитуда. Помимо этих основных ритмов регистрируются и другие ЭЭГ феномены. Например, по мере углубления сна появляются сонные веретена. Это периодическое увеличение частоты и амплитуды тета – ритма. При ожидании команды к действию возникает отрицательная Е – волна ожидания и т.д. В эксперименте ЭЭГ используют для определения уровня активности мозга, а в клинике для диагностики эпилепсии (особенно скрытых форм), а также для выявление смерти мозга (кора живет 3-5 мин., стволовые нейроны 7-10, сердце 90, почки 150). 62. Нейрон, его физиологические свойства, классификация. Особенности возникновения и распространения возбуждения в нейроне. Нейрон – структурно-функциональная единица нервной системы. |