Основы биологической химии предисловие
 Скачать 7.85 Mb. Скачать 7.85 Mb.
|

|
Глава 15. ВЗАИМОСВЯЗЬ ОБМЕНА БЕЛКОВ, ЖИРОВ, УГЛЕВОДОВ И НУКЛЕИНОВЫХ КИСЛОТ Процесс обмена белков, жиров, утлеводов, нуклеиновых кислот тесно взаимосвязан. Эта взаимосвязь осуществляется через ключевые метаболиты, которые служат общим звеном на путях распада или синтеза мономеров. К таким метаболитам можно отнести пируват, ацетил-КоА, фосфоглицерат, метаболиты цикла Кребса (рис. 42). 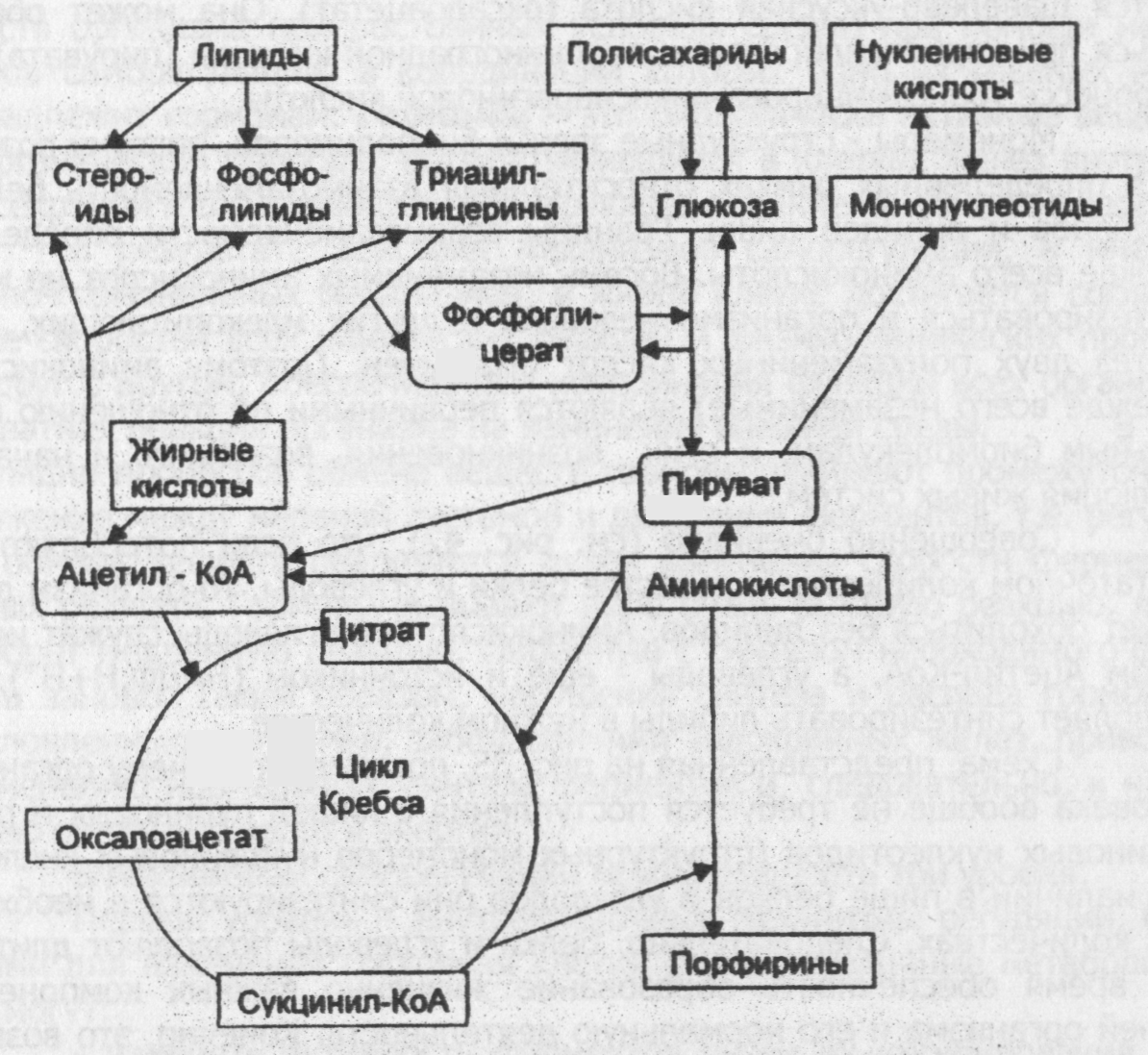 Рис. 42. Взаимосвязь основных путей обмена веществ Из рис. 42 видно, что пируват является точкой пересечения путей распада и синтеза глюкозы (или других моносахаридов) и некоторых аминокислот. Более разветвленный узел метаболических связей представляет собой Ацетил-КоА - через него открывается путь превращения аминокислот (т.е. белков) и глюкозы (т.е. вообще углеводов) в липиды. Как бы вспомогательным связующим звеном между углеводами и липидами служит фосфоглицерат. Через него происходит взаимопревращение углеводов и липидов. Обширные возможности для взаимопревращения одних мономеров в другие представляют промежуточные продукты цикла Кребса. Через цикл Кребса сообщаются все основные пути распада и синтеза веществ. Ацетил-КоА, образующийся в процессе обмена углеводов, жиров и ряда аминокислот, поступает в цикл Кребса, где для его окисления требуется щавелево-уксусная кислота (оксалоацетат). Она может образоваться при карбоксилировании пировиноградной кислоты (пирувата) или в процессе трансаминирования аспарагиновой кислоты. Мономеры - структурные звенья биополимеров, поэтому возможна в определенных рамках относительная взаимозаменяемость белков, углеводов и липидов пищи. Границы взаимозаменяемости определяют прежде всего аминокислоты. Восемь незаменимых аминокислот не могут синтезироваться в организме человека и других млекопитающих, да и синтез двух полузаменимых кислот ограничен. Поэтому аминокислоты (прежде всего незаменимые) являются первичными по отношению к остальным биомолекулам, и с их возникновения, возможно, и началась эволюция живых систем. Совершенно очевидно (см. рис. 42), что если потребляются в достаточном количестве и качестве белки и углеводы, то организм долго может обходиться без липидов. Аминокислоты и углеводы служат источником Ацетил-КоА, а углеводы - еще и источником (НАДФ.Н+Н+), что позволяет синтезировать липиды в нужном количестве. Схема, представленная на рис. 35, показывает, почему организму человека вообще не требуется поступления с пищей пуриновых и пиримидиновых нуклеотидов (структурных мономеров нуклеиновых кислот) - при наличии в пище белков и углеводов они синтезируются в необходимых количествах. Следовательно, белки и углеводы позволяют длительное время обеспечивать образование жизненно важных компонентов тканей организма и его нормальную деятельность. Конечно, это возможно, если с белками и углеводами поступают и такие компоненты пищи, как витамины, вода и минеральные вещества. С другой стороны, человек может долго обходиться без углеводов, если в достаточном количестве поступают остальные компоненты пищи. Таким образом, преобладание распада одних и биосинтеза других пищевых веществ определяется, прежде всего, физиологическим состоянием и потребностями организма в энергии и метаболитах. Глава 16. ГОРМОНЫ, НЕРВНО-ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВЕЩЕСТВ 16.1. Понятие о гормонах. Основные принципы регуляции обмена веществ Одной из уникальных особенностей живых организмов является их слособносгь сохранять постоянство гомеостаза (постоянство многих свойств организма при постоянных условиях среды) при помощи механизмов саморегуляции, в координации которых одно из главных мест принадлежит гормонам. Гормоны - это биологически активные вещества органической природы, вырабатывающиеся в клетках желез внутренней секреции и оказывающие регулирующее влияние на обмен веществ. В результате действия механизмов саморегуляции, а именно нервно-гормональных механизмов, в живой клетке достигается согласование скоростей всех химических реакций и физико-химических процессов друг с другом, обеспечивается координация функций всех органов и адекватная реакция организма на изменения внешней среды. В регуляции процессов обмена веществ гормоны занимают промежуточное положение между нервной системой и действием ферментов, т.е. регуляция обмена веществ реализуется путем изменения скорости ферментативных реакций. Гормоны вызывают либо очень быструю реакцию, либо наоборот медленную реакцию, связанную с синтезом необходимого фермента заново. Таким образом, нарушения синтеза и распада гормонов, обусловленные, например, заболеваниями эндокринных желез, приводят к изменению нормального синтеза ферментов и, следовательно, к нарушению обмена веществ и энергии. В механизмах саморегуляции можно выделить три уровня. Первый уровень - внутриклеточные механизмы регуляции. Сигналами для изменения состояния клетки служат различные метаболиты. Они могут: - изменять активность ферментов путем их ингибирования или активации; - изменять количество ферментов путем регулирования их синтеза и распада; - изменять скорость трансмембранного перекоса веществ. Межорганная координация этого уровня регуляции обеспечивается передачей сигналов двумя путями: через кровь с помощью гормонов (эндокринная система) и через нервную систему. Второй уровень регуляции - эндокринная система. Гормоны освобождаются в кровь на специфический стимул, которым может быть нервный импульс или изменение концентрации какого-то метаболита в крови, протекающей через эндокринную железу (например, снижение концентрации глюкозы). Гормон транспортируется с кровью и, достигая клеток мишеней, модифицирует в них обмен веществ через внутриклеточные механизмы. При этом происходит изменение обмена веществ и устраняется стимул, вызвавший освобождение гормона. Выполнивший свою функцию гормон разрушается специальными ферментами. Третий уровень регуляции - нервная система с рецепторами сигналов как внешней среды, так и внутренней. Сигналы трансформируются в нервный импульс, который в синапсе с клеткой-эффектором вызывает освобождение медиатора - химического сигнала. Медиатор через внутриклеточные механизмы регуляции вызывает изменение обмена веществ. Клетками-эффекторами могут быть и эндокринные клетки, отвечающие на нервный импульс синтезом и выделением гормонов. Все три уровня регуляции тесно взаимосвязаны и действуют как единая нервно-гормональная или нейро-гуморальная система регуляции (рис. 43). Поток информации о состоянии внешней и внутренней среды организма поступает в нервную систему, где перерабатывается, а в ответ посылаются регуляторные сигналы к периферическим органам и тканям. Под прямым контролем нервной системы находятся мозговое вещество надпочечников и гипоталамус. Нервные импульсы, поступающие от различных отделов головного мозга, влияют на секрецию клетками гипоталамуса нейропептидов - либеринов и статинов, регулирующих выделение тропных гормонов гипофиза. Либерины стимулируют синтез и выделение тройных гормонов, а статины - ингибируют. Тройные гормоны гипофиза влияют на секрецию гормонов в периферических железах. Образование и секреция гормонов периферическими железами происходит непрерывно. Это необходимо для поддержания нужного уровня их в крови, так как они быстро инактивируются и выделяются из организма.  Рис. 43. Схема нервно-гормональной регуляции (сплошные стрелки означают синтез гормонов, а пунктирные - влияние гормона на органы-мишени) Концентрация гормонов в крови невелика: порядка 10-6 – 10-11 моль/л. Время полужизни, в основном, несколько минут, для некоторых - десятки минут, очень редко - часы. Требуемый уровень гормона в крови поддерживается за счет механизма саморегуляции по принципу «плюс-минус» межгормональных взаимоотошений. Тропные гормоны стимулируют образование и секрецию гормонов периферическими железами (знак "+"), а последние по механизму отрицательной обратной связи угнетают (знак "-") образование тропных гормонов, действуя через клетки гипофиза (короткая обратная связь) или нейро-секреторные клетки гипоталамуса (длинная обратная связь), рис.44. В последнем случае угнетается секреция либеринов в гипоталамусе. 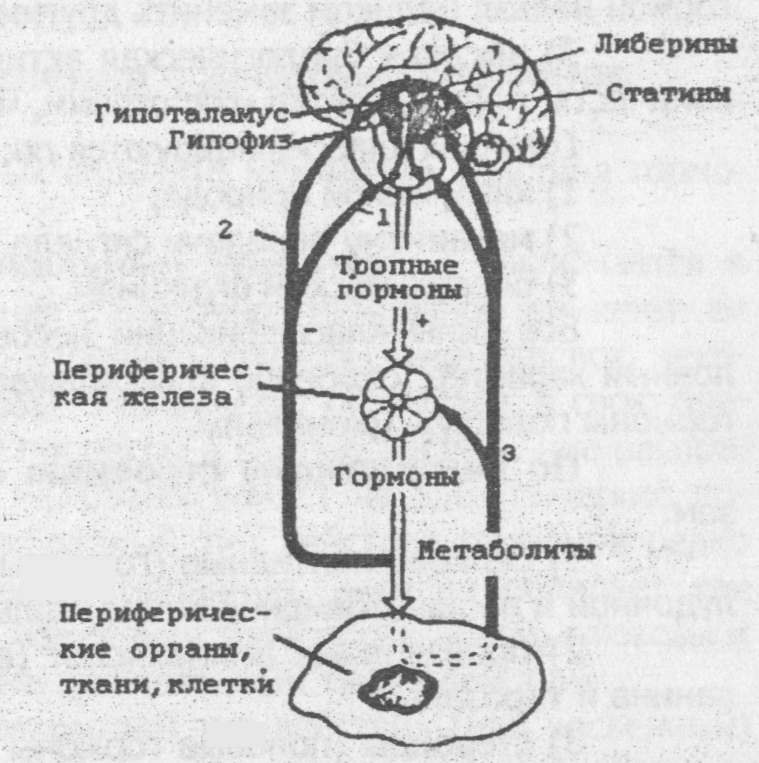 Кроме того существует метаболитно-гормональная обратная связь: гормон, действуя на обмен веществ в тканях, вызывает изменение содержания в крови какого-либо метаболита, а тот по механизму обратной связи влияет на секрецию гормонов в периферических железах или непосредственно (внутриклеточный механизм), или через гипофиз и гипоталамус (см. рис. 44). Такими метаболитами являются глюкоза (индикатор состояния углеводного обмена), аминокислоты (индикатор состояния белкового обмена), нуклеотиды и нуклеозиды (индикаторы состояния нуклеинового и белкового обмена), жирные кислоты, холестерин (индикаторы состояния липидного обмена); Н2О, Са2+, Na+, К+, СI¯ и некоторые другие ионы (индикаторы состояния водно-солевого баланса). 16.2. Классификация гормонов Гормонам присущи следующие общие биологические признаки: 1) дисгантность действия, то есть они регулируют обмен и функции зффекторных клеток на расстоянии; 2) строгая специфичность биологического действия, то есть один гормон нельзя целиком заменить другим; 3) высокая биологическая активность - достаточно очень малых количеств, порой десятка микрограмм, чтобы сохранить жизнь организма. Гормоны классифицируются по: 1) химической природе; 2) механизму передачи сигнала в клетку - мишень; 3) биологическим функциям. Все типы классификации несовершенны и носят несколько условный характер, особенно классификация по функциям, так как многие гормоны полифункциональны. По химическому строению гормоны делят следующим образом: 1) белково-пептидные (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, кальциотонин щитовидной железы); 2) производные аминокислот (адреналин - производное фениланина и тирозина); 3) стероиды (половые гормоны - андрогены, эстрогены и гестагены, кортикостероиды). По биологическим функциям гормоны делят на следующие группы: 1) регулирующие обмен углеводов, жиров, аминокислот - инсулин, глюкагон, адреналин, глюкокортикостероиды (кортизол); 2) регулирующие водно-солевой обмен - минераллокортикостероиды (альдостерон), антидиуретический гормон (вазопрессин); 3) регулирующие обмен кальция и фосфатов - паратгормон, кальцитонин, кальцитриол; 4) регулирующие обмен веществ, связанный с репродуктивной функцией (половые гормоны) - эстрадиол, прогестерон, тестостерон. 5) регулирующие функции эндокринных желез (тройные гормоны) - кортикотропин, тиротропин, гонадотропин. В эту классификацию не включены соматотропин, тироксин и некоторые другие гормоны, которые оказывают полифункциональное действие. Кроме того, помимо гормонов, выделяющихся в кровь и действующих на органы, удаленные от места синтеза гормона, существуют еще гормоны местного действия, регулирующие обмен веществ в тех органах, где они образуются. К ним относят гормоны желудочно-кишечного тракта, гормоны тучных клеток соединительной ткани (гепарин, гистамин), гормоны, выделяемые клетками почек, семенных пузырьков и других органов (простагландины) и т. д. 16.3. Общие представления о действии гормонов Различают три возможных варианта механизма действия гормонов. Мембранный, или локальный, механизм - заключается в том, что гормон в месте связывания с клеточной мембраной изменяет ее проницаемость для метаболитов, например, глюкозы, аминокислот, некоторых ионов. Поступление глюкозы, аминокислот оказывает, в свою очередь, влияние на биохимические процессы в клетке, а изменение распределения ионов по обе стороны мембраны влияет на электрический потенциал и функцию клеток. Мембранный тип действия гормонов редко встречается в изолированном виде. Например, инсулин обладает как мембранным (вызывает местные изменения транспорта ионов, глюкозы и аминокислот), так и мембранно-внутриклеточным типом действия. Мембранно-внутриклеточный тип действия (или косвенный) характерен для гормонов, которые, не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточный химический посредник, который и является полномочным представителем гормона внутри клетки. Гормон через мембранные рецепторы влияет на функцию сигнальных систем (обычно это ферменты), запускающих образование или поступление внутриклеточных посредников. А последние, в свою очередь, влияют на активность и количество разных ферментов и тем самым изменяют обмен веществ в клетке. Цитозольный механизм действия характерен для липофильных гормонов, способных проникать через липидный слой мембраны внутрь клетки, где они вступают в комплекс с цитозольными рецепторами. Этот комплекс регулирует количество ферментов в клетке, избирательно влияя на активность генов хромосом ядра, и тем самым изменяют обмен веществ и функции клетки. Подобный тип действия гормона называется прямым, в отличие от мембранно-внутриклеточного, когда гормон регулирует обмен веществ лишь косвенно, через внутриклеточные посредники. 16.4. Гормоны щитовидной и паращитовидных желез Гормоны щитовидной железы Щитовидная железа секретирует две группы гормонов с разным влиянием на обмен веществ. Первая группа - йодтиронины: тироксин и трийодтиронин. Эти гормоны регулируют энергетический обмен и влияют на деление и дифференцировку клеток, определяя развитие организма. Йодтиронины действуют на многие ткани организма, но в наибольшей степени на ткани печени, сердца, почек, скелетных мышц и в меньшей степени на жировую и нервную ткани. При гиперфункции щитовидной железы (гипертиреозе) наблюдается избыточное образование йодтиронинов. Характерным признаком тиреотоксикоза является ускоренный распад углеводов и жиров (мобилизуются из жировых депо). Быстрое сгорание жирных кислот, глицерина и продуктов гликолиза требует большого расхода кислорода. Митохондрии увеличиваются в размерах, набухают, меняется их форма. Поэтому иногда тиреотоксикоз называют «болезнью митохондрий». Внешне гипертериоз проявляется в виде следующих симптомов: увелечение основного обмена, повышение температуры тела (повышенная теплопродукция), похудание, выраженная тахикардия, повышенная нервная возбудимость, пучеглазие и др. Снимаются эти нарушения или хирургическим удалением части щитовидной железы, или с помощью препаратов, угнетающих ее деятельность. При гипофункции (гипотиреозе) щитовидной железы имеется недостаток йодтиронинов. Гипотериоз в раннем детском возрасте называется кретинизмом или микседемой детей, а у взрослых - просто микседемой. Кретинизм характеризуется выражейной физической и умственной отсталостью. Это объясняется снижением действия йодтиронинов на деление клеток и их дифференцировку, что влечет за собой замедленный и неправильный рост костной ткани, нарушение дифференцировки нейронов. У взрослых микседема проявляется в снижении основного обмена и температуры тела, ухудшении памяти, нарушении кожных покровов (сухость, шелушение) и др. В тканях организма снижен обмен углеводов и жиров и все энергетические процессы. Гипотериоз устраняется лечением препаратами йодтиронинов. Ко второй группе относится кальциотонин (белок с молекулярной массой 30000), он регулирует фосфорно-кальциевый обмен, его действие рассмотренно ниже. Гормоны паращитовидных желез В паращитовидных железах образуются два белковых гормона - кальциотонин (как и в щитовидной железе) и паратгормон (молекулярная масса 3500). Оба гормона регулируют фосфорно-кальциевый обмен: паратгормон повышает уровень кальция и снижает уровень неорганических фосфатов в крови, а кальциотонин снижает содержание в крови и кальция, и фосфатов. Гипофункция желез, или гипопаратериоз, встречается редко и характеризуется повышенной возбудимостью нервно-мышечной системы (судороги). Причина этого заключается в низком содержании ионов Са2+ в крови и межклеточной жидкости, что облегчает деполяризацию мембран, вызываемую током ионов Na+ внутрь клеток, и увеличивает возбудимость нервных и мышечных клеток. Устраняются эти симптомы введением препаратов кальция и паратгормона или витамина D. Гиперфункция, или гиперпаратиреоз, возникает при повышенном образовании паратгормона или его неправильном применении. Симптомами гиперпаратиреоза являются: мобилизация кальция из костей, вплоть до рассасывания отдельных участков костей, что приводит к самопроизвольным переломам. В крови резко повышается содержание Са2+ и снижается содержание фосфора. Кальций начинает оседать во внутренних органах и тканях, что приводит к кальцификации сосудов, почек, желудочно-кишечного тракта и т. д. 16.5. Гормоны поджелудочной железы В тканях островков Лангерганса поджелудочной железы вырабатываются глюкагон, инсулин, соматостатин, а также секретируется гормон местного действия - панкреатический полипептид. Панкреатический полипептид, состоящий из 36 аминокислот, действует на желудочно-кишечный тракт, стимулируя выделение ферментов слизистой желудка и поджелудочной железой. Кроме того, он тормозит перистальтику кишечника и расслабляет желчный пузырь. Соматостатин, первоначально выделенный из гипоталамуса, угнетает выделение соматотропина гипофизом и секрецию глюкагона, гастрина и, возможно, инсулина поджелудочной железой. Уменьшая выделение глюкагона и соматотропина, соматостатин положительно влияет на обмен веществ при сахарном диабете. Глюкагон является белком (молекулярная масса 3485 Да), его секреция усиливается при повышении содержания Са2+ и аргинина в крови и тормозится глюкозой и соматостатином. Глюкагон действует прежде всего на печень, жировую ткань и меньше - на мышцы, вызывая мобилизацию гликогена (гликогенолиз) в печени и отчасти в скелетных мышцах и в жировой ткани. Кроме того, этот гормон тормозит синтез белка в печени и облегчает катаболизм белков до аминокислот, использующихся при образовании мочевины и в глюконеогенезе. В результате повышается в крови содержание глюкозы (следствие гликогенолиза и глюконеогенеза), жирных кислот и глицерина. В целом все происходящие изменения в обмене веществ при действии глюкагона напоминают признаки сахарного диабета. Инсулин, поступающий в кровь, находится в свободной и в связанной с белками плазмы формах. Свободный инсулин влияет на метаболизм всех инсулиночувствительных тканей, а связанный - только на жировую ткань. К инсулиночувствительным тканям относятся мышечная и соединительная (в том числе и жировая) ткани. Секреция инсулина усиливается глюкозой и Са2+, аргинином и лейцином, соматотропином; тормозится соматостатином, но меньше, чем секреция глюкагона. Инсулин действует на ткани через мембранные рецепторы. Комплекс инсулин - рецептор резко изменяет проницаемость клеточных мембран для глюкозы, аминокислот, ионов Са2+, К+, Na+, а именно ускоряет транспорт внутрь клеток глюкозы, аминокислот, ионов К+,Са2+. Инсулин через ферментную систему тормозит гликогенолиз, глюконеогенез, липолиз и облегчает синтез белка. Таким образом, инсулин регулирует углеводный, липидный, белковый и минеральный обмены, по показателям которых судят о действии инсулина. Инсулин снижает концентрацию глюкозы, аминокислот, жирных кислот, глицерина и ионов К+ в крови, а также уменьшает потерю с мочой К+ и аминокислот. В целом действие инсулина анаболитическое. Избыток инсулина (гипогликемия) возникает при опухолях островков Лангерганса или при передозировке инсулина. При этом все изменения обмена веществ, указанные выше, усилены. Следствием этого могут быть обморочные состояния, а в крайнем случае - судороги и смертельный исход. Устраняется, гипогликемия глюкозой и гормономи, вызывающими гипергликемию (глюкагон, адреналин). Дефицит инсулина (гипергликемия) приводит к сахарному диабету - очень широко распространенному заболеванию. При сахарном диабета заметно преобладают катаболические реакции над анаболическими. Взамен глюкозы, которая плохо усваивается в инсулиночувствительных тканях, в организме мобилизуются липиды и усиливается сгорание жирных кислот. В результате в крови увеличивается содержание глюкозы, аминокислот, жирных кислот, глицерина, холестерина, кетоновых тел. Повышенное содержание кетонов вызывает снижение рН крови, что может привести к смертельному исходу. 16.6. Гормоны надпочечников В мозговом веществе надпочечников образуются адреналин и норадреналин. Секреция адреналина усиливается при понижении глюкозы в крови, при стрессах. В целом адреналин действует подобно глюкагону на адреналинозависимые органы-мишени: жировую ткань, скелетные мышцы, печень. Происходят такие же изменения углеводного и липидного обменов, как и под влиянием глюкагона, т.е. усиливается липолиз и гликогенолиз. Адреналин действует также на сердечно-сосудистую систему. Он учащает ритм сердечных сокращений и их силу, повышает кровяное давление, расширяет мелкие артериолы. Кроме того, адреналин расслабляет гладкие мышцы кишечника, бронхов, матки. Адреналин в медицинской практике используется редко: как средство, стимулируёщее сердечные сокращения, повышающее кровяное давление и (вместе с глюкозой) для повышения уровня глюкозы в крови (при передозировке инсулина). В коре надпочечников, из холестерина образуются стероидные гормоны - кортикостероиды, которые делят на три группы: 1) глюкокортикоиды, действующие на углеводный обмен; 2)минералокортикоиды, действующие на минеральный обмен; 3) половые гормоны (мужские-андрогены, женские-эстрогены), выделяющиеся в небольших количествах по сравнению с первыми двумя группами. Действие глюкокортикоидов. Секреция глюкокортикоидов - кортизона (гидрокортизона) и кортикостерона находится под контролем кортикотропина. Выброс кортикотролина из гипофиза является типичным ответом на стресс. Это влечет за собой выделение в кровь глюкокортикоидов, которые, в свою очередь, облегчают выделение адреналина. По механизму отрицательной обратной связи глюкокортикоиды тормозят выделение кортикотропина. Органами-мишенями для глюкокортикоидов являются печень, почки, лимфоидная ткань (селезенка, лимфоузлы, лимфоидные бляшки кишечника, лимфоциты, тимус и др.), соединительная ткань (кости, подкожная соединительная ткань, жировая соединительная ткань и др.), скелетные мышцы. В этих тканях имеются цитозольные рецепторы глюкокортикоидов. Комплекс гормон - рецептор поразному влияет на различные ткани. В печени и почках он усиливает синтез белков, в остальных тканях - наоборот, а в лимфоидной ткани вызывает ее распад (лимфоцитолиз). В результате лимфоцитолиза в кровь поступают аминокислоты, которые используются в печени и почках на синтез белка и в глюконеогенезе. Так как глюкокортикоиды усиливают выделение адреналина из мозгового вещества надпочечников, то тем самым они влияют на обмен жиров, вызывая через адреналин их мобилизацию. Это приводит к повышению в крови содержания глицерина, жирных кислот. Первый используется в глюконеогенезе, а вторые, сгорая в печени, идут на образование кетоновых тел. Таким образом, в результате действия глюкокортикоидов в крови увеличивается концентрация глюкозы, аминокислот, жирных кислот, глицерина, кетонов, т.е. появляются симптомы, аналогичные сахарному диабету. Такой диабет называют «стероидным». В некоторой степени глюкокортикоиды влияют и на водно-минеральный обмен: повышают выделение К* в почках и задерживают Na+ , а с ним и воду во внеклеточных пространствах (что вызывает отеки). Это действие сходно с действием минералокортикоидов, только более слабое. Торможение синтеза белков костной ткани приводит к рассасыванию ее отдельных участков. Кальций и фосфор поступают из них в кровь и теряются с мочой. Глюкокортикоиды широко применяются в практике, основой их лечебного эффекта служит влияние на лимфоидную и соединительную ткани. Лимфоидная ткань участвует в образовании антител в ответ на инфекцию или поступление в организм чужеродных веществ. При повторном контакте с чужеродным веществом происходит взаимодействие его с антителами, что вызывает чрезмерно бурную реакцию организма, которую называют аллергией. Аллергия вызывает воспаление, сопровождающееся местными нарушениями сосудистой проницаемости и повреждением тканей. Разрушенные участки тканей замещаются соединительной тканью, образуются рубцы, деформирующие органы. Глюкокортикоиды, подавляя образование антител в лимфоидной ткани, уменьшают чрезмерную чувствительность организма (сенсибилизацию) к чужеродным веществам, развитие аллергии и воспаления, мешающих нормальной работе органов. Действие минералокортикоидов. Секреция минералокортикоида - альдостерона регулируется ионами К+ и Na+. При низком содержании Na+ или повышенном – К+ в крови усиливается секреция альдостерона. Альдостерон регулирует баланс в организме жизненно необходимых ионов Na+, К+, СI¯ , воды, поэтому без него невозможна нормальная жизнедеятельность организма. Мишенями для альдостерона служат клетки эпителия дистальных канальцев почек, содержащие много циторецепторов для гормона. Комплекс альдостерон - циторецептор действует так, что усиливается реабсорбция Na+ и его противоиона - СI¯ из мочи в межклеточную жидкость и далее в кровь. Одновременно выделяется в мочу К+ (в обмен на Na+) из эпителия канальцев. Таким образом, альдостерон вызывает задержку в организме Na+, СI¯ и воды (вода удерживается Na+ вторично) и потерю с мочой К+. Гиперфункция коры надпочечников (гиперкортицизм) может проявлятся в разных формах. Например, избыточное образование глюкокортикоидов (болезнь Иценко-Кушинга), являющееся следствием поражения гипоталамо-гипофизарной системы, проявляется в виде следующих симптомов: атрофия подкожной соединительной ткани, стероидный диабет, остеопороз (пустоты в костях), гипертония (за счет усиления секреции адреналина и норадреналина). Избыточное образование альдостерона (болезнь Конна) приводит к нарушению водно-солевого обмена. Симптомы: отеки, повышение кровяного давления, повышенная возбудимость миокарда. Если человек употребляет много соли, то развивается так называемая «солевая» гипертония. Гипокортицизм (Аддисонова или бронзовая болезнь) проявляется следующими признаками: снижение устойчивости организма к стрессам и инфекциям, гипогликемия (недостаток глюкокортикоидов); потеря Na+ и воды, накопление К+ в организме, что вызывает гипотонию, мышечную слабость вплоть до бессилия. При гипокортицизме больные погибают из-за нарушений водно-солевого баланса. Половые гормоны рассматриваются ниже. 16.7. Гормоны половых желез Половые железы (гонады) - парные органы, представленные у мужчин семенниками, у женщин - яичниками. Мужские половые гормоны - андрогены, женские - эстрогены. И те, и другие синтезируются из холестерина и в соответствующих количествах имеются и у мужчин, и у женщин. Секреция половых гормонов контролируется гипофизарными гормонами - гонадотропинами: фоллитропином, лютропином и пролактином. По достижении половой зрелости у мужчин выделяются фоллитропин, регулирующий образование сперматозоидов, и лютропин, регулирующий образование андрогенов (тестостерона); тестостерон по механизму отрицательной обратной связи угнетает секрецию лютропина гипофизом. Пролактин у мужчин остается в связанном состоянии в гипофизе и в норме не секретируется. У женщин секретируются все гонадотропины, причем циклически. Это задает тон циклическим процессам в организме женщины, которые называют «половой цикл». Половой цикл включает два тесно связанных и протекающих одновременно процесса: яичниковый цикл (процессы в яичниках) и маточный или менструальный цикл (процессы в матке). Чаще всего продолжительность полового цикла 26-28 дней. Женские половые гормоны - эстрогены (эстрон, эстрадиол, эстриол) обеспечивают протекание следующих физиологических процессов: 1) развитие органов половой сферы (яйцеводов, матки, влагалища), обеспечивающих детородную функцию женщин; 2) формирование вторичных половых признаков в период полового созревания; 3) регуляцию полового цикла; 4) формирование полового инстинкта и психического статуса женщины; 5) протекание беременности и родового акта, развитие молочных желез и подготовку их к лактации в период беременности. Дефицит эстрогенов до периода полового созревания приводит к задержке развития (инфантильности) органов половой сферы, формирования вторичных половых признаков, к нарушению половых циклов. Кроме того, имеет место отрицательный азотистый баланс, потеря кальция и фосфатов. Мужские половые гормоны секретируются под действием гипофизарного гормона - лютропина. Это главным образом тестостерон и его производное - дигидротестостерон. Оба андрогена активные, но первый в большей степени в мышцах, а второй - в простате, семенных пузырьках. Отражением регуляторного действия андрогенов является увеличение биосинтеза белков в тканях, т. е. положительный азотистый баланс организма. Развивается мощная скелетная мускулатура, весь формирующийся в период полового созревания скелет становится более массивным, увеличивается синтез белков в почках и печени, развиваются мужские половые органы и добавочные половые железы (простата, семенные пузырьки), идет развитие вторичных половых признаков (рост волос на лице и теле, формирование характерного мужского тембра голоса). Дефицит андрогенов в организме (евнухоизм) сопровождается недоразвитием половых органов и вторичных половых признаков, отсутствием полового влечения, запаздыванием процессов окостенения эпифизов костей (удлинение конечностей), атрофией скелетной мускулатуры, чрезмерным отложением жира, нарушением корковых процессов торможения. 16.8. Гормоны гипоталамо-гипофизарной системы В передней, и средней доле гипофиза образуются тропные гормоны, задняя доля секретирует только нейрогормоны (вазопрессин и окситоцин). По химическому строению тиреотропин, фоллитропин, лютропин - гликопротеиды. Остальные гормоны - простые белки, имеющие одну полипептидную цепь, а вазопрессин и окситоцин - циклические октапептиды. Секреция тройных гормонов контролируется пептидами гипоталамуса. В настоящее время выделены следующие гипоталамические нейропептиды - регуляторы секреции гормонов гипофиза:
|