Справочный материал. Глава 18 – Эндокринная система. Справочный материал по Физиологии. Глава 18 Эндокринная система
 Скачать 0.66 Mb. Скачать 0.66 Mb.
|

|
Справочный материал по Физиологии. Глава 18 – Эндокринная система. Для эндокринной системы ключевым является понятие «гормон». Гормоны — межклеточные гуморальные химические регуляторы — секретируются во внутреннюю среду организма (преимущественно в кровь) из специализированных (эндокринных) клеток и действуют на клетки–мишени, содержащие молекулы рецепторов к конкретным гормонам. Такое дистантное (через кровоток) взаимодействие между клетками–продуцентами гормонов и клетками–мишенями известно как эндокринная регуляция. Паракринная регуляция подразумевает эффекты гормонов, влияющих путём диффузии на соседние клетки–мишени, а аутокринная — непосредственно на секретирующие эти гормоны клетки (см. рис. 4–10 в книге). Информационные межклеточные взаимодействия, реализуемые гормонами, укладываются в следующую последовательность событий: «сигнал (гормон) рецептор (второй посредник) физиологический ответ». Физиологические концентрации гормонов, осуществляющих гуморальную регуляцию функций, колеблются в пределах 10–7–10–12 М, т.е. гормоны эффективны в крайне низких концентрациях. Разнообразные гормоны и гормональные системы регулируют практически все функции организма, включая метаболизм, репродукцию, рост и развитие, водный и электролитный баланс и поведение. Регуляцию активности многих эндокринных желёз осуществляют при помощи механизмов обратной связи гипофиз и гипоталамус. Синтез некоторых гормонов (адреналин, норадреналин и др.) непосредственно не зависит от регулирующего влияния гипофиза и контролируется симпатической нервной системой. Терминология. Гормоны синтезируются в органах классической эндокринной системы и в клетках так называемой диффузной эндокринной системы. В настоящее время понятие «гормон» значительно расширено: помимо гормонов, вырабатываемых в эндокринной системе, термин «гормон» применяют также по отношению к нейропептидам, интерлейкинам, интерферонам, факторам роста и другим биологически активным веществам, осуществляющим гуморальную (дистантную) регуляцию функций клеток–мишеней. Органы классической эндокринной системы. Термины «гормон», «гормоны», «гормональный» применяют по отношению к органам эндокринной системы и продуцируемым их эндокринными клетками биологически активным веществам — гормонам. К органам классической эндокринной системы относят гипофиз, эпифиз, щитовидную и паращитовидные железы, надпочечник, островки Лангерханса поджелудочной железы, плаценту, яичники и семенники. Диффузная эндокринная система. Под этим термином понимают совокупность отдельных клеток, продуцирующих гормоны и рассыпанных одиночно и мелкими скоплениями в слизистой и подслизистой оболочках трубчатых органов (преимущественно пищеварительной и дыхательной систем). Иногда такие эндокринные клетки называют энтерохромаффинными, аргентаффинными и даже апудоцитами. Этот последний термин (от англ. APUD — Amine Precursor Uptake, Decarboxylase) лучше не применять, так как представление об APUD–системе имеет лишь исторический интерес. Нейропептиды. Термин появился после обнаружения в некоторых нервных клетках биологически активных веществ (но не синаптических нейромедиаторов!), оказывающих регуляторные эффекты на другие клетки. Впоследствии выяснилось, что некоторые из этих нейрогормонов продуцируют и не-нервные клетки, а некоторые нейропептиды выполняют и функции нейромедиаторов. Другие биологически активные вещества. Гуморальную (дистантную) регуляцию выполняют и иные функциональные классы биологически активных веществ (факторы роста, интерлейкины, интерфероны, хемокины, эритропоэтин, предсердный натриуретический фактор, ангиотензины и ряд других), синетезируемых и секретируемых в самых различных органах и клетках. Гормоны, цитокины и хемокины. Хотя термин «гормон» применяют в расширительном смысле (фактически по отношению ко всем биологически активным веществам, секретируемым одними клетками и регулирующим режим функционирования мишеней), наметилась отчётливая тенденция все биологически активные вещества информационного характера подразделять на гормоны, цитокины и хемокины. Согласно этой тенденции, клетки эндокринной системы секретируют гормоны, клетки иммунной системы (и некоторые другие клетки) — цитокины; наконец, хемокины (вещества, оказывающие хемотаксическое действие на мишени) секретируют различные клетки при иммунных реакциях и при воспалении. Химия гормонов По химической структуре гормоны, а также другие биологически активные вещества регуляторного характера (например, факторы роста, интерлейкины, интерфероны, хемокины, ангиотензины, Пг и ряд других) подразделяют на пептидные, стероидные, производные аминокислот и арахидоновой кислоты. Пептидные гормоны относятся к полярным веществам, которые не могут непосредственно проникать через биологические мембраны. Поэтому для их секреции используется механизм экзоцитоза. По этой же причине рецепторы пептидных гормонов встроены в плазматическую мембрану клетки–мишени, а передачу сигнала к внутриклеточным структурам осуществляют вторые посредники. Трансляция мРНК и сборка полипептидной цепочки происходит в цистернах шероховатой эндоплазматической сети (эргастоплазма). Посттрансляционная модификация (например, гликозилирование — присоединение к полипептидной цепочке углеводных молекул, в результате образуются гликопротеины) и формирование секреторных пузырьков происходит в комплексе Гольджи. Экзоцитоз: секреторные мембранные пузырьки, содержащие пептидные гормоны (пептиды, полипептиды, белки, гликопротеины), сливаются изнутри с плазмолеммой секреторной клетки; в результате содержимое пузырьков оказывается снаружи клетки. Экзоцитоз стимулирует вызванное деполяризацией клеточной мембраны увеличение концентрации Ca2+ в цитоплазме. Стероидные гормоны: минералокортикоиды, глюкокортикоиды, андрогены, эстрогены, прогестины, кальцитриол. Эти вещества — производные холестерола — относятся к неполярным, поэтому они свободно проникают через биологические мембраны. По этой причине секреция стероидных гормонов происходит без участия секреторных пузырьков. По этой же причине рецепторы неполярных молекул расположены внутри клетки–мишени. Такие рецепторы в общем виде называют ядерными. Холестерол поступает в клетки извне. Синтез стероидных гормонов — многоэтапный процесс, происходящий при участии десятков ферментов, расположенных в гладкой эндоплазматической сети и в митохондриях. Производные аминокислот — тирозина (йодсодержащие гормоны щитовидной железы, норадреналин, адреналин и дофамин), гистидина (гистамин), триптофана (мелатонин и серотонин) Производные тирозина Неполярные молекулы тироксина (T4) и трийодтиронина (Т3) образуются путём их отщепления от йодированного (зрелого) тиреоглобулина в фаголизосомах тиреоцитов и через плазмолемму базальной части тиреоцитов поступают в кровь. Их рецепторы относятся к ядерным. Катехоламины не проникают через мембраны, они накапливаются в секреторных пузырьках и секретируются из клеток путём экзоцитоза. Их рецепторы встроены в плазматическую мембрану клеток–мишеней. Производное гистидина — гистамин [4-(2-аминоэтил)-имидазол] — продукт декарбоксилирования гистидина. Рецепторы гистамина встроены в плазматическую мембрану клеток–мишеней. Производные триптофана — мелатонин (N ацетил-5метокситриптамин) и серотонин (5гидрокситриптамин). Их рецепторы встроены в плазматическую мембрану клеток–мишеней. Производные арахидоновой кислоты (эйкозаноиды, или простаноиды). Эйкозаноиды (от греч. eikosi — двадцать) состоят (как и арахидоновая кислота) из 20 атомов углерода. К ним относятся простагландины (Пг), тромбоксаны, простациклины, лейкотриены, гидроксиэйкозотетраеноевая (HETE, от англ. hydroxyeicosatetraenoic) и эпоксиэйкозотриеноевая кислоты, а также производные этих кислот. Все эйкозаноиды обладают высокой и разносторонней физиологической активностью, многие из них функционируют только внутри клетки. Арахидоновая кислота — жирная кислота, мобилизуемая из фосфолипидов клеточной мембраны фосфолипазой A2. Арахидоновая кислота образуется также при расщеплении диацилглицерола. Арахидоновая кислота окисляется трояко: с помощью циклооксигеназ (циклооксигеназный путь, образуются Пг, тромбоксаны, простациклины), липооксигеназы (липооксигеназный путь, образуются лейкотриены, 5-HETE) и эпоксигеназы арахидоновой кислоты (CYP2J2 из семейства цитохромов P450, образуются HETE и эпоксиэйкозотриеноевая кислота). Простагландины. Известны ПгG2 (предшественник ПгH2), ПгH2 (предшественник простациклинов и тромбоксанов), ПгI2 (его иногда называют простациклином), ПгD2, ПгE2 и ПгF2. Эти биологически активные эндогенные алифатические кислоты регулируют множество функций: увеличивают проницаемость сосудистой стенки, влияют на сократимость ГМК сосудов и бронхов, изменяют порог болевой чувствительности, стимулируют секрецию желудочного сока. Пг применяют для стимуляции родов. Развитие ряда патологических состояний также связано с действием Пг (воспаление, бронхиальная астма, рост опухолей), а PGE2 является мощным пирогеном и модулятором метастазирования раковых клеток. Простациклины (в том числе ПгI2) образуются из ПгI2, ингибируют агрегацию тромбоцитов; вызывают расслабление ГМК кровеносных сосудов. Тромбоксаны — группа соединений, влияющих на агрегацию тромбоцитов, вызывают также сокращение ГМК кровеносных сосудов. Лейкотриены. Известны лейкотриены A4 (предшественник остальных лейкотриенов) B4, C4, D4, E4, F4. Лейкотриены влияют на активность многих ионных каналов (непосредственно или посредством протеинкиназ) и на процессы экзоцитоза, вызывают сокращение ГМК кишечника и кровеносных сосудов, являются медиаторами воспаления. HETE — гидроксиэйкозотетраеноевые кислоты и их производные способствуют выбросу Ca2+ из внутриклеточных кальциевых депо в моноцитах и макрофагах, являются сильными вазоконстрикторами, способствуют пролиферации разных типов клеток. Эпоксиэйкозотриеноевые кислоты стимулируют выброс Ca2+ из внутриклеточных кальциевых депо, стимулируют трансмембранный Na+-H+–обмен, пролиферацию клеток, являются вазодилататором. Из мембранных фосфолипидов также образуется фактор активации тромбоцитов (PAF), относящийся к наиболее сильным спазмогенам. Механизмы действия гормонов на клетки–мишени Информационные межклеточные взаимодействия, реализуемые в эндокринной системе, предусматривают такую последовательность событий: гормон рецептор клетки–мишени (второй посредник) ответ клетки–мишени Каждый гормон оказывает на клетку–мишень регулирующий эффект тогда и только тогда, когда он в качестве лиганда связывается со специфичным для него белком–рецептором в составе клетки–мишени. Циркуляция в крови. Гормоны циркулируют в крови либо свободно, либо в комплексе со связывающими их белками (T4, Т3, стероидные гормоны, инсулиноподобные факторы роста, гормон роста). Связывание с такими белками существенно увеличивает время полужизни гормонов. Так, T4 в составе комплекса циркулирует около 1–й нед, тогда как время полужизни свободного T4 составляет несколько минут. Рецепторы и вторые посредники рассмотрены в главе 4 «Межклеточные взаимодействия». Гормоны и их физиологические эффекты В этом разделе дана физиологическая характеристика различных гормонов, синтезируемых и секретируемых клетками эндокринной системы. Гипоталамо-гипофизарная система Часть промежуточного мозга — гипоталамус — и отходящий от его основания гипофиз анатомически и функционально составляют единое целое — гипоталамо-гипофизарную эндокринную систему (рис. 18–1, 18–2, 18–3, см. также рис. 16–3,В,Г). 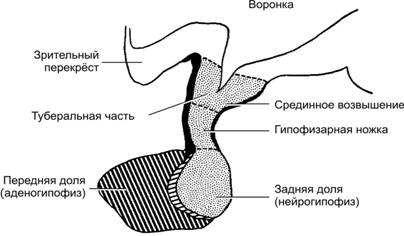 Рис. 18–1. Анатомические взаимоотношения гипоталамуса и ножки гипофиза [11]. Точечная штриховка: срединное возвышение и задняя доля гипофиза (нейрогипофиз) имеют нейральное происхождение и фактически являются частью гипоталамуса. Косая штриховка: эпителиальная часть гипофиза (аденогипофиз) развивается из эктодермы ротовой бухты. Роль гипоталамо–гипофизарной системы для эндокринной регуляции функций организма столь велика, что её иногда называют «президентом эндокринного общества». Гипоталамус В нейросекреторных нейронах гипоталамуса синтезируются нейропептиды, поступающие как в переднюю (рилизинг–гормоны), так и в заднюю (окситоцин и вазопрессин) доли гипофиза. Рилизинг–гормоны Гипоталамические рилизинг–гормоны (от англ. releasing hormone) — группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза. С функциональной точки зрения рилизинг–гормоны подразделяют на либерины (рилизинг–гормоны, способствующие усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза) и статины (рилизинг гормоны, подавляющие синтез и секрецию гормонов в клетках–мишенях). К гипоталамическим либеринам относятся соматолиберин, гонадолиберин, тиреолиберин и кортиколиберин, а статины представлены соматостатином и пролактиностатином. Соматостатин — циклический тетрадекапептид C76H104N18O19S2 (мол. масса 1637,9) — синтезируется многими нейронами ЦНС (в том числе нейронами околожелудочковой области гипоталамуса), клетками островков Лангерханса поджелудочной железы, эндокринными клетками ЖКТ и ряда других внутренних органов. Функции. Соматостатин — мощный регулятор функций эндокринной и нервной систем, ингибирует синтез и секрецию множества гормонов и секретов. Передняя доля гипофиза. Соматостатин подавляет синтез и секрецию гормона роста, АКТГ и тиреотропного гормона. Островки Лангерханса. Соматостатин подавляет секрецию инсулина и глюкагона. Энтероэндокринная система. Соматостатин ингибирует секрецию гастрина, холецистокинина, секретина. Почка. Соматостатин подавляет секрецию ренина. Желудок. Соматостатин ингибирует желудочную секрецию. Рецепторы. Соматостатин реализует эффекты через связанные с G белком мембранные высокоаффинные рецепторы (ингибирование аденилатциклазы цАМФ ... уменьшение [Ca2+] в цитозоле блокада экзоцитоза секреторных пузырьков). Идентифицировано не менее 5 подтипов рецепторов соматостатина, имеющих выраженную органную специфичность распределения. Соматолиберин (соматокринин) — пептид, содержащий 44 аминокислотных остатка, синтезируется нейросекреторными нейронами дугового ядра (n. arcuatus) гипоталамуса и некоторыми опухолями островковых клеток поджелудочной железы (соматолибриномы). Функция. Гипоталамический соматолиберин стимулирует секрецию гормона роста в передней доле гипофиза. Рецептор соматолиберина относят к семейству связанных с G белком рецепторов типа секретина (стимуляция аденилатциклазы цАМФ протеинкиназа A активация кальциевых каналов типа L увеличение [Ca2+] в цитозоле экзоцитоз секреторных пузырьков). Гонадолиберин (люлиберин) и пролактиностатин. Ген LHRH кодирует аминокислотные последовательности для гонадолиберина и пролактиностатина. Мишени гонадолиберина — гонадотрофы, а пролактиностатина — лактотрофы передней доли гипофиза. Функции. Гонадолиберин — ключевой нейрорегулятор репродуктивной функции, стимулирует синтез и секрецию ФСГ и ЛГ в продуцирующих гонадотрофы клетках, а пролактиностатин подавляет секрецию пролактина из лактотрофных клеток передней доли гипофиза. Рецепторы люлиберина — трансмембранные гликопротеины, связанные с G белком. Активирование G белка приводит к образованию инозитолтрифосфата и диацилглицерола, что в конечном итоге стимулирует секрецию ФСГ и ЛГ. Тиреолиберин — трипептид, синтезируется многими нейронами ЦНС (в том числе нейросекреторными нейронами паравентрикулярного ядра). Мишени тиреолиберина — тиреотрофы и лактотрофы передней доли гипофиза. Пролактин. Тиреолиберин стимулирует секрецию пролактина из лактотрофов. Тиреотропный гормон. Тиреолиберин стимулирует секрецию тиреотропина из тиреотрофов. СТГ. В патологических ситуациях тиреолиберин стимулирует секрецию гормона роста из соматотрофов. Рецепторы. Связывание тиреолиберина с рецепторами активирует в клетках–мишенях G белок, что приводит к активации фосфолипазы A2 и синтезу производных арахидоновой кислоты. Кортиколиберин — пептид из 41 аминокислотного остатка — синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах. Влияние глюкокортикоидов на синтез кортиколиберина Гипоталамус. Глюкокортикоиды подавляют синтез гипоталамического кортиколиберина. Плацента. Глюкокортикоиды стимулируют синтез плацентарного кортиколиберина. Функции кортиколиберина многочисленны. Передняя доля гипофиза. Стимуляция синтеза и секреции АКТГ и других продуктов экспрессии гена проопиомеланокортина. Стресс. Кортиколиберин — координатор эндокринных, нейровегетативных и поведенческих ответов в стрессовых ситуациях. Последние всегда затрагивают ось «кора головного мозга–гипоталамус–гипофиз–надпочечник». Плацента. На сроке 18 нед беременности по содержанию в крови кортиколиберина можно выделить группы беременных с преждевременными и поздними родами. К предсказанному сроку содержание в крови кортиколиберина резко возрастает. Возможно, этот гормон является сигналом начала родов. Рецепторы кортиколиберина относят к семейству связанных с G белком рецепторов типа секретина (стимуляция аденилатциклазы цАМФ протеинкиназа A активация кальциевых каналов типа L увеличение [Ca2+] в цитозоле экзоцитоз секреторных пузырьков). Меланостатин — L-пролил-L-лейцилглицинамид — подавляет образование меланотропинов. Либерины и статины по аксонам гипоталамических нейронов достигают срединного возвышения, где секретируются в кровеносные сосуды портальной системы кровотока (рис. 18–2), далее по воротным венам гипофиза эти нейрогормоны поступают в переднюю долю гипофиза и регулируют активность её эндокринных клеток (табл. 18-1, см. также рис. 18–4). 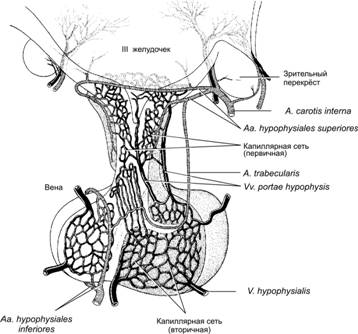 |