экзамен биология. Сущность жизни. Основные уровни организации и свойства живого. Жизнь
 Скачать 0.92 Mb. Скачать 0.92 Mb.
|

|
Транскрипция Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает «инструкция» о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией. Синтез мРНК начинается с обнаружения РНК-полимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции — промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплементарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5'-конца к 3'-концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3'-концом (3' → 5'). Такую цепь называют кодогенной. Антипараллельность соединения двух полинуклеотидных цепей в молекуле ДНК позволяет РНК-полимеразе правильно выбрать матрицу для синтеза мРНК. Продвигаясь вдоль кодогенной цепи ДНК, РНК-полимераза осуществляет постепенное точное переписывание информации до тех пор, пока она не встречает специфическую нуклеотидную последовательность — терминатор транскрипции. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и от вновь синтезированной мРНК. Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор, образует единицу транскрипции — транскриптон. В процессе синтеза, по мере продвижения РНК-полимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции мРНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов мРНК, шифрующие аминокислоты, называют кодонами. Последовательность кодонов мРНК шифрует последовательность аминокислот в пептидной цепи. Кодонам мРНК соответствуют определенные аминокислоты Матрицей для транскрипции мРНК служит кодогенная цепь ДНК, обращенная к ферменту своим 3'-концом синтеза РНК в области терминатора, отделение полимеразы от завершенной цепи РНК. Процессинг Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК, — альтернативный сплайсинг. Полиаденилирование Полиаденилирование заключается в присоединении к 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты, осуществляемом специальным ферментом poly(A)-полимераза. Сплайсинг После полиаденилирования мРНК подвергается удалению интронов. Процесс катализируется сплайсосомой и называется сплайсингом. Биосинтез белка Молекулы белков по существу представляют собой полипептидные цепочки, составленные из отдельных аминокислот. Но аминокислоты недостаточно активны, чтобы соединиться между собой самостоятельно. Поэтому, прежде чем соединиться друг с другом и образовать молекулу белка, аминокислоты должны активироваться. Эта активация происходит под действием особых ферментов. Причем каждая аминокислота имеет свой, специфически настроенный на нее фермент. Источником энергии для этого (как и для многих процессов в клетке) служит аденозинтрифосфат (АТФ). Важным является то, что каждой аминокислоте соответствует строго специфическая т-РНК. Она находит «свою» аминокислоту и переносит ее в рибосому. Поэтому такая РНК и получила название транспортной. В рибосому поступают различные активированные аминокислоты, соединенные со своими т-РНК. Рибосома представляет собой как бы конвейер для сборки цепочки белка из поступающих в него различных аминокислот. Одновременно с т-РНК, на которой «сидит» своя аминокислота, в рибосому поступает «сигнал» от ДНК, которая содержится в ядре. В соответствии с этим сигналом в рибосоме синтезируется тот или иной белок, тот или иной фермент (так как ферменты являются белками). Синтезированная и-РНК поступает в рибосому и как бы передает этой структуре план — в каком порядке должны соединяться друг с другом поступившие в рибосому активированные аминокислоты, чтобы синтезировался определенный белок. Иначе, генетическая информация, закодированная в ДНК, передается на и-РНК и далее на белок. Транспортная РНК со своей аминокислотой подходит к определенному кодону и-РНК и соединяется с ним; к следующему, соседнему участку и-РНК присоединяется другая т-РНК с другой аминокислотой и так далее, до тех пор пока не будет считана вся цепочка и-РНК и пока не нанижутся все аминокислоты в соответствующем порядке, образуя молекулу белка. А т-РНК, которая доставила аминокислоту к определенному участку полипептидной цепи, освобождается от своей аминокислоты и выходит из рибосомы. Затем снова в цитоплазме к ней может присоединиться нужная аминокислота, и она снова перенесет ее в рибосому. В процессе синтеза белка участвует одновременно не одна, а несколько рибосом — полирибосомы. У организмов, обладающих настоящим ядром (животные, растения), транскрипция и трансляция строго разделены в пространстве и времени: синтез различных РНК происходит в ядре, после чего молекулы РНК должны покинуть пределы ядра, пройдя через ядерную мембрану Затем в цитоплазме РНК транспортируются к месту синтеза белка— рибосомам. Лишь после этого наступает следующий этап — трансляция. У бактерий, ядерное вещество которых не отделено от цитоплазмы мембраной, транскрипция и трансляция идут одновременно Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. Свойства генетического кода Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже). Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными. Строение плазмолеммы: надмембранный, мембранный и субмембранный комплексы. Медицинские аспекты функций плазмолеммы. Клетки многоклеточных организмов, как животных, так и растительных, обособлены от своего окружения оболочкой. Плазмолемма, животных клеток образована мембраной, покрытой снаружи слоем гликокаликса толщиной 10—20 нм. Основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Изнутри к мембране примыкает кортикальный (корковый) слой цитоплазмы толщиной 0,1—0,5 мкм, в котором не встречаются рибосомы и пузырьки, но в значительном количестве находятся микротрубочки и микрофиламенты, имеющие в своем составе сократимые белки. Благодаря свойству избирательной проницаемости она регулирует химический состав внутренней среды клетки. В плазмолемме размещены молекулы рецепторов, которые избирательно распознают определенные биологически активные вещества (гормоны). В удержании этих веществ на клеточной поверхности участвуют белки кортикального слоя. Наличие в оболочке рецепторов дает клеткам возможность воспринимать сигналы извне, чтобы целесообразно реагировать на изменения в окружающей их среде или состоянии организма. В пластах и слоях соседние клетки удерживаются благодаря наличию разного вида контактов, которые представлены участками плазмолеммы, имеющими особое строение. Плазмолемма выполняет следующие функции: 1. Ограничение и обособление клеток. Обособление клеток от межклеточной среды обеспечивается плазматической мембраной, защищающей клетки от механического и химического воздействий. Плазматическая мембрана обеспечивает также сохранение разности концентраций метаболитов и неорганических ионов между внутриклеточной и внешней средой. 2. Контролируемый транспорт веществ и ионов определяет внутреннюю среду клетки и обеспечивает подержание внутриклеточного гомеостаза. 3. Восприятие внеклеточных сигналов (рецепторная функция) и их передача внутрь клетки. 4. Контактное взаимодействие с межклеточным матриксом и взаимодействие с другими клетками при слиянии клеток и образовании тканей. 6. Соединение с элементами цитоскелета обеспечивает поддержание формы клеток и движение гиалоплазмы внутри клетки. Классификация органоидов клетки по строению и по функции. Строение и функции мембранных органоидов клетки: митохондрии, эндоплазматическая сеть, пластинчатый комплекс, пероксисомы. Митохондрии — это структуры округлой или палочковидной, нередко ветвящейся формы Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матрикс органеллы. В нем обнаруживаются зерна. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген. В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала — этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки. Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме. Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата —АТФ). В целом этот процесс называется окислительным фосфорилированием. В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая). Канальцевая и вакуолярная системы образованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяют шероховатую и гладкую цитоплазматическую сети. Эндоплазматическая сеть (ЭПС)–это система цистерн и трубочек, связанных между собой в единое внутриклеточное пространство, отграниченное от остальной части цитоплазмы замкнутой внутриклеточной мембраной. Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называются эргастоплазмой. Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечнополосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения. Пластинчатый комплекс Голъджи образован совокупностью диктиосом Диктиосома представлена стопкой из 3—12 уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки. Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности, пероксисомы. Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. Немембранные органоиды клетки: рибосомы, филаменты, центриоли, реснички, жгутики. Цитоскелет клетки. К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран Рибосома — это округлая рибонуклеопротеиновая частица диаметром 20—30 нм. Состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. На полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока). Микротрубочки - трубчатые образования различной длины. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов. Микрофиламенты - длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов. Актиновые микрофиламенты благодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы. По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10 нм — промежуточные филстенты. В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Выполняют, по-видимому, механическую, каркасную функцию. Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации. Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр, в состав которого входят центриоли. Центриолъ (под электронным микроскопом) имеет вид «полого» цилиндра. Ее стенка образована 27 микротрубочками, сгруппированными в 9 триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза. Организация наследственного аппарата клеток человека. Морфофункциональная характеристика и классификация хромосом. Методы изучения кариотипа человека. Хромосомы – высокая степень конденсации хроматина, постоянно присутствующего в клеточном ядре. В хромосомах сосредоточена большая часть наследственной информации. В основу идентификации хромосом положены следующие признаки: Общая длина хромосомы Размещение центромеры Вторичная перетяжка и др. Различают четыре типа строения хромосом: Телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); Акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом); Субметацентрические (с плечами неравной длины, напоминающие по форме букву L); Метацентрические (V-образные хромосомы, обладающие плечами равной длины). Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.  Расположение центромеры определяет три основных типа хромосом: 1) равноплечие – с плечами равной или почти равной длины; 2) неравноплечие, имеющие плечи неравной длины; 3) палочковидные – с одним длинным и вторым очень коротким, иногда с трудом обнаруживаемым плечом. хромосомный набор-Кариоти́п - совокупность признаков полного набора хромосом, присущая клеткам данного биологического вида, данного организма или линии клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора. Термин «кариотип» был введён в 1924 году советским цитологом. Гигантские хромосомы Такие хромосомы, для которых характерны огромные размеры, можно наблюдать в некоторых клетках на определённых стадиях клеточного цикла. Например, они обнаруживаются в клетках некоторых тканей личинок двукрылых насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щёток). Именно на препаратах гигантских хромосом удалось выявить признаки активности генов. Политенные хромосомы Впервые обнаружены Бальбиани в 1881-го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз, кишечника, трахей, жирового тела и мальпигиевых сосудов личинок двукрылых. Бактериальные хромосомы Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе парасексуального процесса. Кариотип человека (от греч. - орех, ядро и - отпечаток, тип) - диплоидный хромосомный набор человека, представляющий собой совокупность морфологически обособленных хромосом, внесённых родителями при оплодотворении. Хромосомы набора генетически неравноценны: каждая хромосома содержит группу разных генов. Все хромосомы в кариотипе человека делятся на аутосомы и половые хромосомы. В кариотипе человека 44 аутосомы (двойной набор) - 22 пары гомологичных хромосом и одна пара половых хромосом - XX у женщин и ХУ у мужчин. Методы изучения кариотипа человека: Генеалогический метод — составление родословного дерева многих поколений и изучение типа наследования (доминантный или рецессивный, сцепленный с полом или аутосомный), частоты и интенсивности проявления наследственных свойств. Результатом изучения обычно является определение типа наследования, а также риска проявления наследственных нарушений у потомков; Цитогенетический метод — изучение хромосомных наборов здоровых и больных людей. Результат изучения — определение количества, формы, строения хромосом, особенности хромосомных наборов обоих полов, а также хромосомных нарушений; Биохимический метод — изучение изменений в биологических параметрах организма, связанных с изменением генотипа. Результат изучения — определение нарушений в составе крови, в околоплодной жидкости и т. д.; Близнецовый метод — изучение генотипических и фенотипических особенностей однояйцевых и разнояйцевых близнецов. Результат изучения — определение относительного значения наследственности и окружающей среды в формировании и развитии человеческого организма; Популяционный метод — изучение частоты встречаемости аллелей и хромосомных нарушений в популяциях человека. Результат изучения — определение распространения мутаций и естественного отбора в популяциях человека. Поток информации в клетке. Схема реализации наследственной информации. Поток информации в клетке осуществляется в двух направлениях: Транскрипция трансляция полипептид Репликация ДНК деление материнской клетки и образование дочерних(митоз) В потоке информации участвуют ядро (ДНК хромосом, иРНК, переносящие информацию в цитоплазму), аппарат трансляции (рибосомы,тРНК,ферменты,регуляторные белки),геномы митохондрий и хлоропластов. Реализация генетической информации- процесс, происходящий внутри каждой клетки, во время которого генетическая информация ,записанная в ДНК, воплощается в биологически активных веществах- РНК и белках. Основные стадии процесса реализации генетической информации. Начальная стадия хранения информации. После окончания клеточного деления, хроматин, который содержит ДНК с генетической информацией находится в так называемом конденсированном состоянии, которое предназначено для того, чтобы в наиболее сохранном виде доставить генетическую информацию из родительской клетки в дочерние. В этом состоянии ДНК находится в максимально компактном состоянии и не работает. Деконденсания хроматина. Когда деление завершено, ДНК должна быть приведена в активизированное состояние. Для этого она разворачивается под управлением специальных белков хроматина. На этой стадии происходит процесс индукции или ҫугорессии тех или иных генов, когда они могут становиться либо «говорящими» (экспрессируются), либо «молчащими». Одним из проявлений этого процесса является дифференциация клеток. Транскрипция (переписывание) К развёрнутым участкам ДНК получают доступ специальные ферменты, называемые РНК-полимеразами. ДНК и РНК представляет собой цепочку из звеньев — нуклеотидов. Между нуклеотидами ДНК и РНК существует химическое сродство, что позволяет полимеразе двигаться по ДНК и синтезировать РНК, в точности соответствующую ДНК. Полученная в результате транскрипции РНК называется информационной (ИРНК) или матричной (МРHК). Трансляция и транспорт аминокислот. Основными органическими веществами всех живых организмов на Земле являются белки, а в основе всех белков лежит двадцать аминокислот, Каждый белок представляет собой цепочку из аминокислотных молекул. Чтобы «прочитать» информацию из созданных на предыдущем этапе МPHK, требуется во- первых, постоянная подача аминокислот, а во-вторых, работа по преобразованию генетического кода в аминокислотный. Дело в том, что каждой аминокислоте соответствует тройка нуклеотидов и это соответствие в достаточной мере произвольно. Поэтому в клетке всегда присутствует 20 видов так называемых транспортных РНК (ТРНК), которые с одного конца имеют химическое сродство к некоторой тройке нуклеотидов, а с другого конца специальным ферментом (аминоацил-ТРНК-синтетаза) присоединяется соответствующая данной тройке аминокислота. То есть, каждая такая ТРНК является адаптором, а набор молекул синтетазы, которых тоже 20 видов-таблицей преобразования генетического кода в аминокислотный. ТРНК постоянно «вылавливают» плавающие в цитоплазме клетки аминокислоты и доставляют их к месту синтеза белков-к рибосомам. Синтез (сборка) белков в рибосомах. Рибосомы плавают в цитоллазме клетки и к ним поступают МРНК с информацией из ядра и тРНК с материалом из окружающей цитоплазмы. Рибосома также похожа на застежку-молнию, только гораздо крупнее PHК-полимеразы и представляет собой целую клеточную органеллу. Во время работы она надевается на целочку МРHК и скользит по ней. Поступающие в рибосому TРНК соединяются с текущим участком MPHК только в том случае, если ответная часть соответствует закодированной аминокислоте. После этого рибосома получает нужную аминокислоту, отсоединяет её от ТРНК и подсоединяет к белковой цепочке, которую она ткет. Свободная ТРНК удаляется, а рибосома переходит к следующей тройке нуклеотидов, после чего процесс повторяется. Оканчивается он тогда, когда будет пройдена вся цепочка MРHК, при этом будет соткан в точности тот белок, который был закодирован в том гене в ДНК, который дал начало всему процессу. Биологическая активность белков. Белки являются практически универсальными химическими соединениями. Различные аминокислоты обладают различным химическим сродством друг к другу, к воде, к другим веществам. Поэтому, готовый белок скручивается в пространстве особым, только ему присущим образом, обнажая свои части, различные форме и химической активности. В клетках белки используются и как строительный материал и как ферменты, катализирующие или ингибирующие необходимые реакции. Каждый белок можно считать «молекулярным роботом», выполняющим свою примитивную функцию. В результате активности белков, в клетке синтезируются все другие химические вещества, небелкового происхождения, в частности, сами аминокислоты, нуклеотиды, ДНК и РНК и другие. Морфофункциональная и генетическая характеристика стадий митоза. Патологические митозы, их значение. Эндомитоз. Понятие о стволовых клетках. Важным компонентом клеточного цикла является митотический (пролиферативный) цикл —комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткой многоклеточного организма специфических функций, а также периоды покоя. В периоды покоя ближайшая судьба клетки не определена: она может либо начать подготовку к митозу, либо приступить к специализации в определенном функциональном направлении. Длительность цикла регулируется путем изменения продолжительности всех его периодов. Биологическое значение митотического цикла состоит в том, что он обеспечивает преемственность хромосом в ряду клеточных поколений, образование клеток, равноценных по объему и содержанию наследственной информации. Таким образом, цикл является всеобщим механизмом воспроизведения клеточной организации эукариотического типа в индивидуальном развитии. Главные события митотического цикла заключаются в редупликации (самоудвоении) наследственного материала материнской клетки и в равномерном распределении этого материала между дочерними клетками. Указанным событиям сопутствуют закономерные изменения химической и морфологической организации хромосом — ядерных структур, в которых сосредоточено более 90% генетического материала эукариотической клетки (основная часть внеядерной ДНК животной клетки находится в митохондриях). Хромосомы во взаимодействии с внехромосомными механизмами обеспечивают: а) хранение генетической информации, б) использование этой информации для создания и поддержания клеточной организации, в) регуляцию считывания наследственной информации, г) удвоение (самокопирование) генетического материала, д) передачу его от материнской клетки дочерним. По двум главным событиям митотического цикла в нем выделяют репродуктивную и разделительную фазы, соответствующие интерфазе и митозу классической цитологии. В начальный отрезок интерфазы (постмитотический, пресинтетический, или Gi-период) восстанавливаются черты организации интерфазной клетки, завершается формирование ядрышка, начавшееся еще в телофазе. Из цитоплазмы в ядро поступает значительное (до 90%) количество белка. В цитоплазме параллельно реорганизации ультраструктуры интенсифицируется синтез белка. Это способствует росту массы клетки. Если дочерней клетке предстоит вступить в следующий митотический цикл, синтезы приобретают направленный характер: образуются химические предшественники ДНК, ферменты, катализирующие реакцию редупликации ДНК, синтезируется белок, начинающий эту реакцию. Таким образом осуществляются процессы подготовки следующего периода интерфазы — синтетического. В синтетическом или S-периоде удваивается количество наследственного материала клетки. За малыми исключениями редупликация ДНК осуществляется полуконсервативным способом. Он заключается в расхождении биспирали ДНК на две цепи с последующим синтезом возле каждой из них комплементарной цепочки. В результате возникают две идентичные биспирали. Молекулы ДНК, комплементарные материнским, образуются отдельными фрагментами по длине хромосомы, причем неодномоментно (асинхронно) в разных участках одной хромосомы, а также в разных хромосомах. Затем участки (единицы репликации — репликоны) новообразованной ДНК «сшиваются» в одну макромолекулу. Число их меняется в онтогенезе. Смысл редупликации ДНК репликонами становится понятным из следующих сопоставлений. Участки хромосом, в которых начинается синтез, называют точками инициации. Возможно, ими являются места прикрепления интерфазных хромосом к внутренней мембране ядерной оболочки. Можно думать, что ДНК отдельных фракций, о которых речь пойдет ниже, редуплицируется в строго определенной фазе S-периода. Так, большая часть генов рРНК удваивает ДНК в начале периода. Редупликация запускается поступающим в ядро из цитоплазмы сигналом, природа которого не выяснена. Синтезу ДНК в репликоне предшествует синтез РНК. В клетке, прошедшей S-период интерфазы, хромосомы содержат удвоенное количество генетического материала. Наряду с ДНК в синтетическом периоде интенсивно образуются РНК и белок, а количество гистонов строго удваивается. Незначительная часть митохондриальной ДНК редуплицируется в синтетическом, тогда как основная —в постсинтетическом периоде интерфазы. Отрезок времени от окончания синтетического периода до начала митоза занимает постсинтетический (предмитотический), или G2-neриод интерфазы. Он характеризуется интенсивным синтезом РНК и особенно белка. Завершается удвоение массы цитоплазмы по сравнению с началом интерфазы. Это необходимо для вступления клетки в митоз. Часть образуемых белков (тубулины) используется в дальнейшем для построения микротрубочек веретена деления. Синтетический и постсинтетический периоды связаны с митозом непосредственно. Это позволяет выделить их в особый период интерфазы — препрофазу.
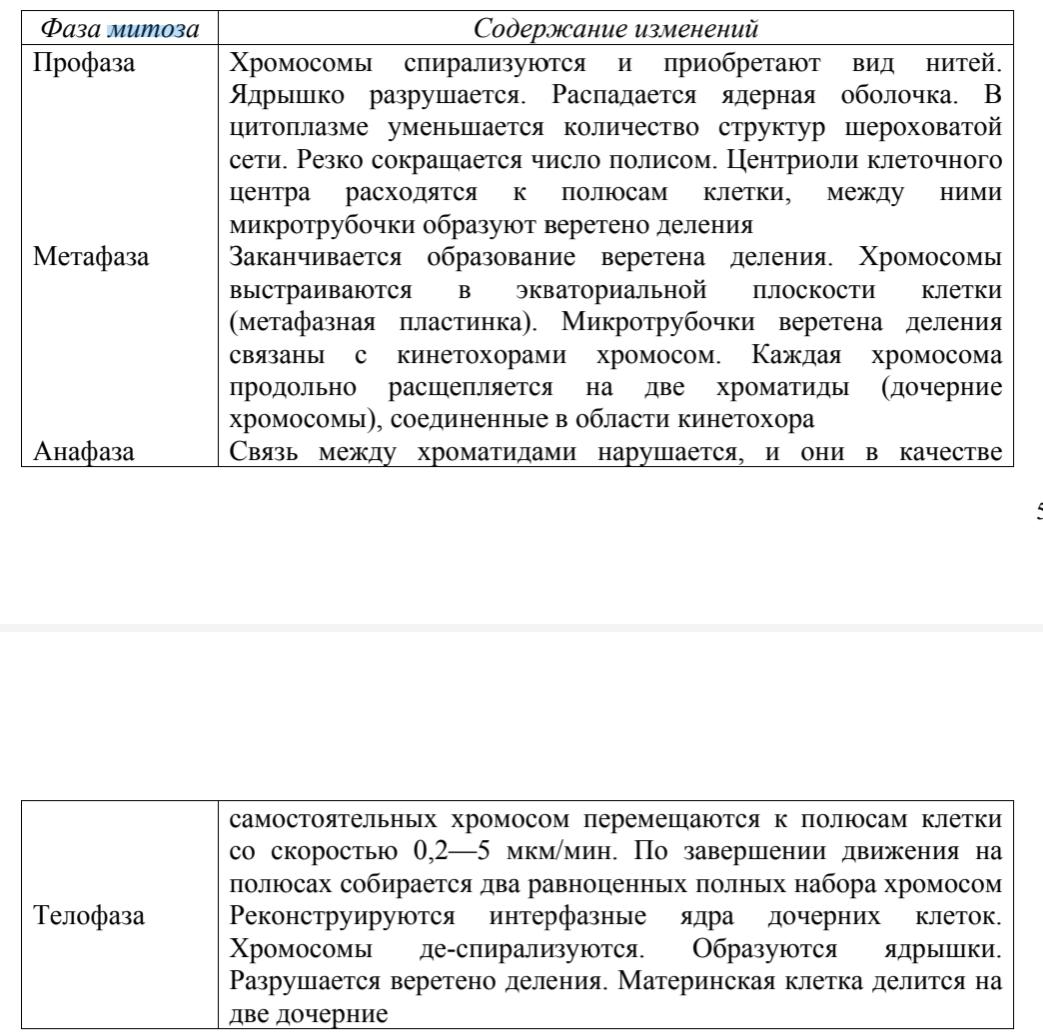 митозе можно выделить четыре фазы. Главные события по отдельным фазам представ Наряду с преобразованием строения и упорядоченными перемещениями хромосом обязательным для разделительной фазы цикла является построение митотического аппарата. Он состоит из системы микротрубочек (ахроматиновое веретено, или веретено деления светооптической микроскопии) и структур, поляризующих митоз1, т.е. обозначающих два полюса в клетке, к которым разойдутся дочерние хромосомы. В клетках млекопитающих и человека роль поляризующих структур выполняют центриоли. Митотический аппарат обеспечивает направленное перемещение дочерних хромосом в анафазе. Для этого необходим контакт микротрубочек со специализированными участками хромосом — центромерами (кинетохорами). При разрушении центромер в эксперименте расхождения хромосом к полюсам клетки не происходит. Природа сил, обеспечивающих расхождение, неизвестна. Предполагают, что функциональную основу митотического аппарата составляет механохимическая система, сходная с действующей в поперечно-полосатой мышце. В нее входят сократимые белки и фермент, катализирующий расщепление АТФ для обеспечения процесса энергией. Нарушения той или иной фазы митоза приводят к патологическим изменениям клеток. Отклонение от нормального течения процесса спирализации может привести к набуханию и слипанию хромосом. Иногда наблюдается отрыв участка хромосомы, который, если он лишен центромеры, не участвует в анафазном перемещении к полюсам и теряется. Отставать при движении могут отдельные хроматиды, что приводит к образованию дочерних ядер с несбалансированными хромосомными наборами. Повреждения со стороны веретена деления приводят к задержке митоза в метафазе, рассеиванию хромосом. При изменении количества центриолей возникают многополюсные или асимметричные митозы. Нарушение цитотомии приводит к появлению дву- и многоядерных клеток. Данные генетики и цитологии указывают на сохранение структурной индивидуальности хромосом в клеточном цикле. Есть свидетельства в пользу упорядоченного размещения хромосом в объеме интерфазного ядра. Особенности взаиморасположения хромосом могут иметь большое функциональное значение. Так, пространственная близость в клетках человека хромосом 13, 14, 15, 21 и 22-й пар, содержащих гены рРНК, объясняется, по-видимому, их участием в формировании ядрышка и образовании рибосом, а 11-й и 16-й хромосом —в образовании молекулы гемоглобина. Есть указания на то, что в клетках разных типов расположение хромосом не одинаково. На основе митотического цикла возник ряд механизмов, с помощью которых в том или ином органе количество генетического материала и, следовательно, интенсивность обмена могут быть увеличены при сохранении постоянства числа клеток. Удвоение ДНК клетки не всегда сопровождается ее разделением на две. Поскольку механизм такого удвоения совпадает с предмитотической редупликацией ДНК и оно сопровождается кратным увеличением количества хромосом, это явление получило название эндомитоза. С генетической точки зрения, эндомитоз — геномная соматическая мутация, о чем будет сказано ниже. Другое явление, сходное по результату и названное политенией, заключается в кратном увеличении содержания ДНК в хромосомах при сохранении их диплоидного количества. Эндомитоз и политения приводят к образованию полиплоидных клеток, отличающихся кратным увеличением объема наследственного материала. В таких клетках в отличие от диплоидных гены повторены более чем два раза. Пропорционально увеличению числа генов растет масса клетки, что повышает ее функциональные возможности. В организме млекопитающих полиплоидизация с возрастом свойственна печеночным клеткам. Стволовые клетки – это клетки-предшественники всех клеток и тканей нашего организма. Стволовые клетки способны поддерживать свою численность с помощью деления и обладают способностью дифференцироваться (превращаться) в различные типы клеток. Гаметогенез. Характеристика периодов, отличия ово- и сперматогенеза. Особенности гаметогенеза человека. Регуляция гаметогенеза и его возможные нарушения. Гаметогенез-образование зрелых половых клеток. Выделяют: Сперматогенез- образование мужских половых клеток Овогенез - образование женских половых клеток. Гаметы - это высокоспециализированные клетки, которые выполняют функцию генеративную. Сперматозоид состоит из трех частей: головки, шейки и жгутика. на передней части головки располагается акросома с хеморецепторами, там же имеются вакуоли с ферментами. внутри головки находится ядро и центриоль. в шейке находится множество митохондрий. Яйцеклетка крупная неподвижная, с большим количеством зернистой цитоплазмы. имеет две оболочки - белковую и желточную. В гаметогенезе выделяют три стадии. Стадия размножения, Стадия роста Стадия созревания В сперматогенезе четвертая - стадия формирования. Сперматогенез у человека начинается после полового созревания. весь этот процесс занимает примерно70 суток. В стадию размножения делятся клетки зачаткового эпителия и образуются многочисленные сперматогонии. Деление в данном случае - митоз. Стадия роста. Соответствует первой интерфазе первого деления мейоза. Меняется характеристика клетки (2н4с) - образуются сперматоциты первого порядка. Стадия созревания. соответствует двум делениям мейоза. Получаются гаплоидные клетки - сперматоциты второго порядка (н2с). далее - второе деление. получаются сперматиды (нс). Стадия формирования. Идет дифференцировка. К большинства мужчин гаметогенез не прекращается до смерти. Изменяется только количество сперматозоидов. Овогенез начинается в эмбриональном периоде жизни. Стадия размножения. Первичные половые клетки делятся путем митоза и образуются овогонии (2н2с). Стадия роста. Появляются овоциты первого порядка(2н4с). Стадия созревания. Овоцит первого порядка начинает мейоз, но первое деление прерывается на стадии профазы-1 до полового созревания. После полового созревания первое деление продолжается. Заканчивается и появляется овоцит второго порядка и мелкая клетка (редукционное тельце первое) (н2с). Затем идет овуляция. Начинается второе деление (в маточных трубах). Оно идет до стадии метафазы-2. Овоцит второго порядка имеет только одну центриоль. Вторая центриоль появляется от сперматозоида. После этого второе деление заканчивается. Выделяется второе редукционное тельце. Редукционное тельце забирает с собой лишние хромосомные наборы. обычно их получается три. Различия сперматогенеза и овогенеза: Разное число стадий Число зрелых половых клеток, образующихся на одну исходную Продолжительность процесса, начало процесса Непрерывность процесса (сперматогенез непрерывен, овогенез имеет две паузы) Производительность (сперматозоидов очень много, не менее 500 млрд., а яйцеклеток 300-400). Нарушение гаметогенеза Гаметогенез протекает в половых железах, которые являются гормонозависимыми органами. Необходимы половые гормоны и два гормона гипофиза - фолликулостимулирующий и лютеинизирующий. Первый у женщин способствует развитию овоцитов и синтезу эстрогенов. У мужчин способствует созреванию сперматозоидов. Второй у женщин способствует развитию желтого тела после овуляции и синтезу гестагенов. у мужчин стимулируют выработку тестостеронов. Нарушения гаметогенеза связаны с нарушением гормонального статуса организма (опухоли, эндокринные заболевания), при хроническом стрессе, привычных интоксикации. Нарушения гаметогенеза, например нерасхождение отдельных пар хромосом, могут приводить к возникновению хромосомно аномальных гамет. Формы осеменения организмов. Оплодотворение, фазы оплодотворения. Биологическая сущность оплодотворения. Искусственное осеменение и оплодотворение, их использование в медицине. Процессу оплодотворения (слияние ядер мужской и женской гамет) предшествует осеменение. Осеменение — процессы, обуславливающие встречу сперматозоида и яйцеклетки. Взаимодействие гамет обеспечивается выделением особых веществ — гамонов (гиногамонов и андрогамонов). Гиногамон I стимулирует подвижность сперматозоида. Гиногамон II блокирует двигательную активность сперматозоидов и способствует их фиксации на оболочке яйцеклетки. Андрогамон I тормозит движение сперматозоидов, что предохраняет их от преждевременной растраты энергии. Андрогамон II способствует растворению оболочки яйцеклетки. Существует 2 способа осеменения: Наружное Внутреннее. У некоторых животных наблюдается кожное осеменение, которое является переходной формой. Это характерно для немертин, пиявок. Этапы оплодотворения: Сближение гамет, акросомная реакция и проникновение сперматозоида; Активация яйца, его синтетических процессов; Слияние гамет (сингамия). Наружная фаза. Сближение гамет относится к наружной фазе. Женские и мужские гаметы выделяют специфические соединения, которые называются гамонами. Яйцеклетками продуцируются гиногамоны I и II, сперматозоидами — андрогамоны I и II. Гиногамоны I активизируют движение сперматозоидов и обеспечивают контакт с яйцом, а андрогамоны II растворяют оболочку яйца. В момент контакта сперматозоида с наружной оболочкой яйца начинается акросомная реакция. Из акросомы выделяется фермент гиалуронидаза. В месте контакта сперматозоида с плазматической мембраной яйца образуется выпячивание или бугорок оплодотворения. Бугорок оплодотворения способствует втягиванию сперматозоида внутрь яйца. Мембраны гамет сливаются. Слияние мужских и женских половых клеток называется сингамия. В ряде случаев (у млекопитающих) сперматозоид проникает в яйцо без активного участия бугорка оплодотворения. Ядро и центриоль сперматозоида переходят в цитоплазму яйца, что способствует завершению мейоза II в ооците. Внутренняя фаза. Характеризуется кортикальной реакцией со стороны яйцеклетки. Происходит отслойка желточной оболочки, которая затвердевает и называется оболочкой оплодотворения. В момент завершения мейоза формируется мужской и женский пронуклеусы. Оба пронуклеуса сливаются. Слияние ядер гамет — синкариогамия составляет сущность процесса оплодотворения, в результате чего образуется зигота. |