Гистология реферат. Тема 1 клетка Вопрос Химический состав, организация плазмолеммы. Функции плазмолеммы
 Скачать 3.29 Mb. Скачать 3.29 Mb.
|

|
Тема 20: «Мужская половая система» Вопрос 1. Развитие мужской половой системы. Первичные гоноциты, начальная локализация, пути миграции в зачаток гонады. Половая дифференцировка. Источниками половых желёз являются урогенитальные валики и первичные половые клетки. Урогенитальные (гонадные) валики — индифферентные гонады — зачатки будущих половых желёз (яичек и яичников). На 4-й неделе эмбриогенеза в грудопоясничном отделе нефротома (на медиальной стороне мезонефроса) формируются гонадные валики, не идентифицируемые ни как мужские, ни как женские половые железы. Индифферентные гонады состоят из заселяемого первичными половыми клетками коркового и мозгового вещества. 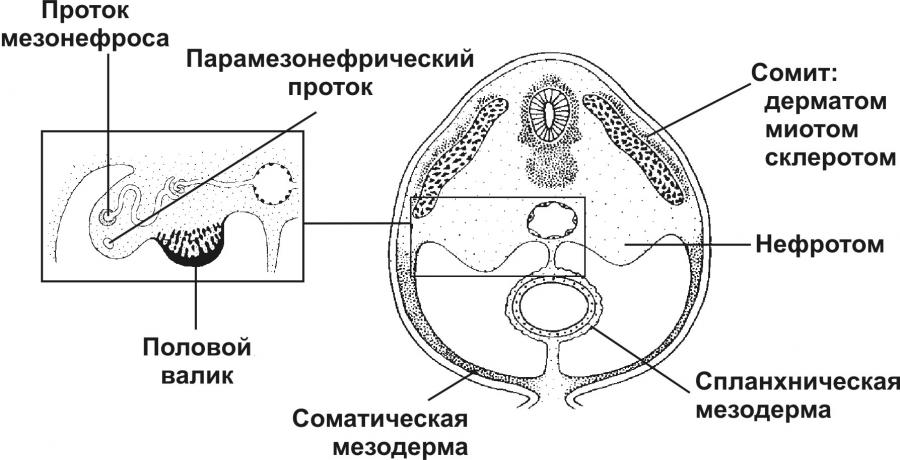 Первичные половые клетки возникают, на 2-й неделе эмбрионального развития из клеток головного отдела эпибласта. В ходе гаструляции первичные половые клетки через первичную полоску в энтодерму желточного мешка и далее в гонадные валики. В плодном периоде первичные половые клетки дифференцируются в овогонии в развивающихся яичниках или в сперматогонии в яичках. На пути от овогоний или сперматогоний до зрелых гамет различают несколько стадий: размножения, роста, созревания и формирования. ПОЛОВАЯ ДИФФЕРЕНЦИРОВКА Хромосомная детерминация пола происходит при оплодотворении, Y‑хромосома — потенциальная детерминанта генетически мужского пола Фактор, детерминирующий развитие мужских гонад (TDF) — один из индукторов развития мужской половой железы. Регуляторный фактор TDF, кодируемый геном Y-хромосомы SRY (Sex-determining Region Y), ответственен за дифференцировку яичек из изначально бипотентных зачатков гонад. Индифферентные гонады Критическая стадия развития индифферентных гонад — 8-я неделя внутриутробного развития. До 45–50 дня зачатки гонад не имеют половой дифференцировки. Под влиянием фактора транскрипции TDF гонадные валики развиваются как яички; при отсутствии эффектов этого фактора развиваются яичники. Дифференцировку других структур определяют мужские половые гормоны и мюллеров ингибирующий фактор (MIF ¾ Mullerian Inhibiting Factor), продуцируемые в яичках плода. Дифференцировка по мужскому типу. При кариотипе 46XY из клеток мезенхимы гонадного валика дифференцируются интерстициальные эндокринные клетки яичек плода. Под контролем гонадотропинов (хорионического и гипофизарного) интерстициальные эндокринные клетки яичек плода секретируют тестостерон. Экспрессия гена SRY в поддерживающих эпителиальных клетках инициирует транскрипцию гена, кодирующего MIF. Дифференцировка по женскому типу при кариотипе 46ХХ происходит при отсутствии определяющего развитие яичек фактора Y-хромосомы, андрогенов и MIF. Гонадные валики развиваются как яичники автономно, под влиянием гормонов плаценты. Яичник и яичко Дифференцировка яичника и яичка из индифферентных гонад приведена на рисунке. В женском организме в индифферентных гонадах развивается преимущественно корковое и атрофируется мозговое вещество. В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме. Вопрос 2. Яичко. Развитие и строение. Канальцы яичка. В мужском организме преимущественное развитие получает мозговое вещество индифферентной гонады. На 8-й неделе эмбриогенеза яички располагаются на уровне верхних поясничных позвонков. От нижнего полюса яичка вниз тянется поддерживающая связка, выполняющая функцию проводника яичка из брюшной полости через паховый канал в мошонку. Опущение яичек в мошонку завершается приблизительно за 1 месяц до рождения и не происходит при крипторхизме. Яички (testis), в отличие от яичников, располагаются вне полости тела (в мошонке). Это обстоятельство важно для нормального течения сперматогенеза, происходящего при температуре 34 °C и ниже. Снаружи яичко покрыто соединительнотканной белочной оболочкой (t. albuginea). Внутренний её слой богат кровеносными сосудами — сосудистая оболочка (t. vasculosa). Утолщение белочной оболочки, вдающееся с одной стороны в паренхиму яичка, называется средостением (mediastinum testis). От средостения внутрь яичка отходят перегородки, разделяющие его на дольки конической формы. Каждая долька содержит от одного до четырёх извитых семенных канальцев (tubulus seminifer contortus), выстланных сперматогенным эпителием (epithelium spermatogenicum). В рыхлой соединительной ткани между семенными канальцами расположены интерстициальные эндокринные клетки (endocrinocytus interstitialis). По мере приближения к средостению семенные канальцы продолжаются в прямые (tubuli recti). Их стенка образована кубическим эпителием и базальной мембраной, окружённой тонкой соединительнотканной оболочкой. Прямые канальцы впадают в сеть яичка (rete testis) — систему анастомозирующих тонкостенных трубочек, продолжающихся в выносящие канальцы придатка. Эпителий, выстилающий сеть яичка, представлен кубическими клетками. Некоторые клетки на апикальной поверхности имеют по одной ресничке, способствующей продвижению сперматозоидов. Под базальной мембраной эпителия располагается рыхлая соединительная ткань, окружающая трубочки rete testis снаружи. Генеративная функция (сперматогенез, spermatogenesis) осуществляется в извитых семенных канальцах, а эндокринная (синтез мужских половых гормонов) — в интерстициальных эндокринных клетках.  Яичко с придатком. А — соединительнотканная строма яичка и придатка. Б — паренхима яичка и придатка. Яичко (кратко) Функции: Сперматогенез; гаметы(22X, 22Y) Эндокринная функция(тестостерон, эстрогены, ингибин, мюллеров ингибирующий фактор, андроген связывающий белок) Строение: Медиальная, латеральная и передняя поверхность яичка покрыты брюшиной(tunica vaginalis) Белочная оболочка (tunica albuginea)плотная соединительнотканная полностью окружающая яичко Сосудистая оболочка (tunica vasculosa) Соединительнотканные трабекулы делят паренхиму органа на 250долек, содержащих 1-4 извитых семенных канальцев Средостение содержит сеть яичка (rete testis) Интерстициальная соединительная ткань (между канальцами) содержит эндокринные клетки Лейдига. Вопрос 3. Извитые семенные канальцы, строение стенки. Клеточный состав сперматогенного эпителия. Извитые семенные канальцы выстланы сперматогенным эпителием, содержащим клетки двух типов — гаметы с их предшественниками на различных стадиях дифференцировки (сперматогонии, сперматоциты первого порядка, сперматоциты второго порядка, сперматиды, сперматозоиды) и поддерживающие эпителиальные клетки (сустентоциты). Снаружи канальцы окружены тонкой соединительнотканной оболочкой. Извитые семенные канальцы открываются в прямые, по которым сперматозоиды попадают в сеть яичка. 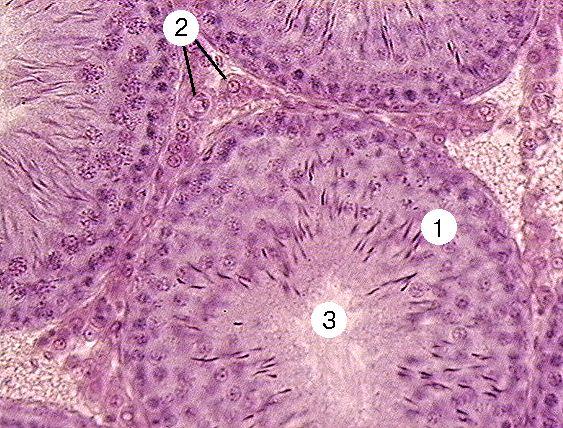 Извитые семенные канальцы. Канальцы выстланы сперматогенным эпителием (1). Между канальцами видны крупные интерстициальные эндокринные клетки (2). В просвете канальцев (3) находятся сперматозоиды. Окраска гематоксилином и эозином. Сустентоциты Широкое основание сустентоцитов (sustentocytus, поддерживающих эпителиальных клеток) находится на базальной мембране, а суженная складчатая апикальная часть достигает просвета канальца. При помощи плотных контактов сустентоциты делят сперматогенный эпителий на базальное и адлюминальное пространства. В базальном пространстве находятся только сперматогонии. В адлюминальном пространстве располагаются сперматоциты первого и второго порядков, сперматиды и сперматозоиды (см. рис. 15-12). Между сустентоцитами и гаметами устанавливаются адгезионные и щелевые контакты (рис. 15-13). Сустентоциты оказывают физическую поддержку развивающимся гаметам, обеспечивают их питательными веществами (трофическая функция), поглощают продукты метаболизма, фагоцитируют остатки цитоплазмы формирующихся сперматозоидов (резидуальные тельца) и дегенерирующие половые клетки, секретируют жидкость для транспорта сперматозоидов в семенных канальцах. Гематотестикулярный барьер. Поддерживающие клетки в период полового созревания вблизи базальной мембраны образуют плотные контакты, формирующие гематотестикулярный барьер. Благодаря такому барьеру, в адлюминальном пространстве создаётся специфическая гормональная среда с высоким уровнем тестостерона. Барьер изолирует созревающие половые клетки от токсических веществ и препятствует развитию аутоиммунного ответа против поверхностных Аг, экспрессирующихся на мембране созревающих сперматозоидов. Сустентоциты связаны плотными, адгезионными и щелевыми контактами. Между поддерживающими эпителиальными клетками в их базальной части локализуются плотные контакты, отвечающие за формирование гематотестикулярного барьера. В апикальной части клетки связаны щелевыми контактами. В плотных контактах молекулы адгезионного контакта JAM, окклюдины и клаудины через ZO-1 и ZO-2 взаимодействуют с актином. Между сустентоцитами и гаметами устанавливаются адгезионные и щелевые контакты. Продукты секреции. В кровь сустентоциты секретируют эстрогены, ингибин, мюллеров ингибирующий фактор MIF (в плодном периоде). В адлюминальном пространстве поддерживающие эпителиальные клетки накапливают андроген-связывающий белок, фактор стволовых клеток, трансферрин, активаторы плазминогена. · Андрогенсвязывающий белок — фактор, обеспечивающий поддержание высокой концентрации тестостерона в сперматогенном эпителии путём накопления гормона в просвете семенных канальцев. · Фактор стволовых клеток (SCF) необходим для сохранения популяции сперматогоний. · Трансферрин, кроме транспорта железа в сперматогенный эпителий, является мощным митогенным фактором. · Протеазы (катепсины, активаторы плазминогена) и ингибиторы протеаз (серпины, цистатины) влияют на протеолитические реакции, что важно для миграции созревающих половых клеток из базального пространства в адлюминальное. Фолликулостимулирующий гормон (фоллитропин) — главный стимулирующий фактор сустентоцитов. Рецепторы фолликулостимулирующего гормона, относятся к связанным с G–белком, активирующим аденилатциклазу. Увеличение в цитозоле цАМФ повышает секреторную функцию поддерживающих эпителиальных клеток. Вопрос 4. Сперматогенез. Характеристика его основных фаз. Гормональная регуляция сперматогенеза. Сперматогенез (путь от сперматогонии до сперматозоида) в извитых семенных канальцах длится 65 дней, но окончательная дифференцировка сперматозоидов происходит в протоке придатка яичка в течение следующих двух недель. Только в области хвоста придатка сперматозоиды становятся зрелыми половыми клетками и приобретают способность к самостоятельному передвижению и оплодотворению яйцеклетки. Сперматогенез подразделяют на стадии размножения, роста, созревания и формирования. На стадии размножения выделяют сперматогонии типов А и В (рис. 15-12). Сперматогонии типа А Среди сперматогоний типа А по степени конденсации хроматина различают тёмные и светлые клетки. Тёмные сперматогонии считаются резервными стволовыми клетками, редко вступающими в митоз. Светлые сперматогонии — полустволовые клетки, находящиеся в непрерывно следующих друг за другом клеточных циклах (интерфаза сменяется митозом). В результате деления светлой клетки типа А образуются либо две клетки типа В (симметричный митоз), либо одна клетка типа В и одна светлая клетка типа А (асимметричный митоз). Сперматогонии типа В Сперматогонии типа В имеют круглое ядро и конденсированный хроматин. Они также вступают в митоз, но при этом остаются связанными друг с другом при помощи цитоплазматических мостиков. После ряда митотических делений сперматогонии типа В дифференцируются в сперматоциты первого порядка, которые из базального пространства перемещается в адлюминальное и вступают в стадию роста. В стадии роста объём сперматоцитов первого порядка увеличивается в 4 и более раз. Клеточные ассоциации На стадиях размножения, роста и созревания сперматогенные клетки входят в состав клеточных ассоциаций. Светлая сперматогония типа А формирует клон сперматогенных клеток (синцитий), в котором клетки остаются связанными цитоплазматическими мостиками до стадии формирования. Клеточная ассоциация в своем развитии от сперматогоний до сперматозоидов проходит шесть стадий (I–VI), для каждой из которых существует характерный тип сочетания сперматогенных клеток. В ходе сперматогенеза клеточные ассоциации находятся на различных стадиях развития, что обусловливает мозаичное распределение клеточных ассоциаций в составе сперматогенного эпителия. 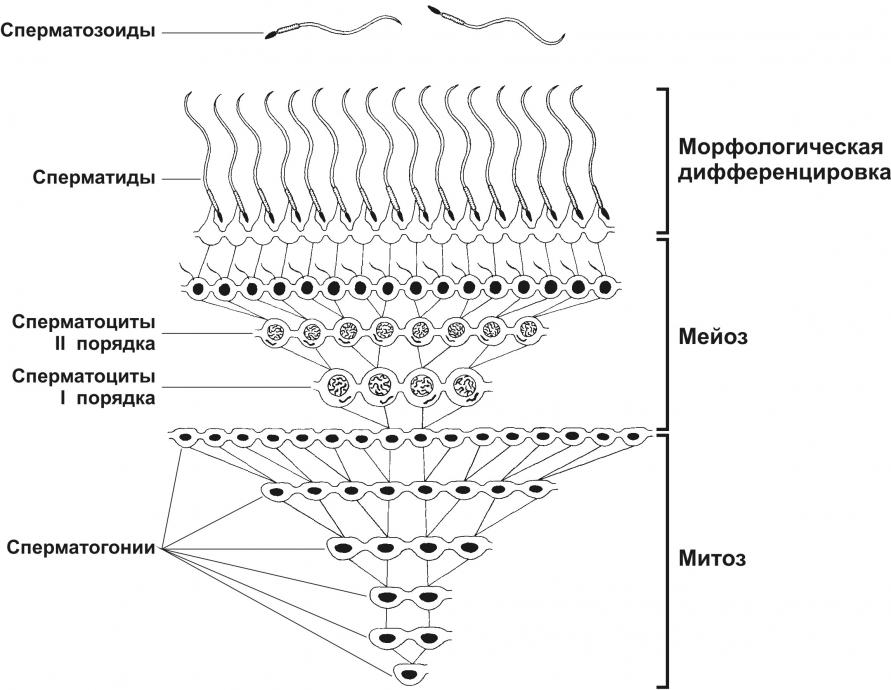 Клон сперматогенных клеток. На стадиях размножения, роста и созревания сперматогенные клетки связаны цитоплазматическими мостиками. Только на стадии формирования гамет цитоплазматические мостики разрываются, и сперматозоиды оказываются свободными. Созревание За стадией роста наступает стадия созревания, которая состоит из следующих друг за другом двух делений мейоза. В результате первого деления из одного сперматоцита первого порядка образуется два сперматоцита второго порядка, а после второго деления — четыре сперматиды. Сперматиды располагаются вблизи просвета канальца. Каждая сперматида вчетверо мельче сперматоцита первого порядка и имеет ядро с гаплоидным набором хромосом. Формирование Спермиогенез — постмейотическая стадия морфологической дифференцировки сперматиды в сперматозоид. В составе комплекса Гольджи (на одном из полюсов ядра) появляется акросомный пузырёк с акросомной гранулой, одновременно центриоли мигрируют на противоположную сторону ядра и инициируют сборку аксонемы. Акросомная гранула заполняет акросомный пузырёк и образуется акросомная шапочка. Хроматин конденсируется, ядро вытягивается и приобретает окончательную форму. Цитоплазматические микротрубочки, растущие от центриолей, образуют цилиндрическую манжетку, способствующую элонгации сперматиды. Митохондрии располагаются спиралеобразно вокруг аксонемы. Сперматида переворачивается хвостом в просвет канальца. Между клетками разрываются цитоплазматические мостики, а сформировавшиеся сперматозоиды оказываются свободными. Пластичность межклеточных контактов в сперматогенном эпителии. Выход сперматоцитов из базального пространства в адлюминальное и последующее отделение дифференцированных сперматозоидов от поддерживающих клеток обусловлены дезинтеграцией межклеточных контактов как между сустентоцитами, так между сустентоцитами и гаметами. Главную роль в дезинтеграции контактов играют протезы и цитокины, в т.ч. TGFb 3 и TNFa. Высокие концентрации тестостерона и Ca2+ способствуют восстановлению межклеточных контактов. Вопрос 5. Спермиогенез. Спермиогенез: Постмейотическая стадия морфологических изменений сперматид с образованием сперматозоидов Фаза Гольджи: 1 В составе комплекса Гольджи появляется акросомный пузырёк с акросомной гранулой. 2 Центриоли мигрируют на противоположную сторону клетки и инициируют сборку аксонемы Фаза головной шапочки: Акросомный пузырёк уплощается, покрывает 2/3 поверхности ядра Фаза акросомы: 1.Акросомная гранула заполняет акросомный пузырёк → акросома 2.Хроматин конденсируется, ядро вытягивается 3.Микротрубочки образуют цилиндрическую манжетку 4.Митохондрии мигрируют и спиралеобразно располагаются вокруг аксонемы 5.Сперматида переворачивается хвостом в просвет канальца Фаза созревания: 1.Ядро приобретает окончательную форму 2.Сперматида избавляется от лишней цитоплазмы, разрушается манжетка 3.Разрываются цитоплазматические мостики. Вопрос 6. Строение и функции сустентоцитов (клеток Сертоли). Сустентоциты Широкое основание сустентоцитов (sustentocytus, поддерживающих эпителиальных клеток) находится на базальной мембране, а суженная складчатая апикальная часть достигает просвета канальца. При помощи плотных контактов сустентоциты делят сперматогенный эпителий на базальное и адлюминальное пространства. В базальном пространстве находятся только сперматогонии. В адлюминальном пространстве располагаются сперматоциты первого и второго порядков, сперматиды и сперматозоиды Между сустентоцитами и гаметами устанавливаются адгезионные и щелевые контакты Сустентоциты оказывают физическую поддержку развивающимся гаметам, обеспечивают их питательными веществами (трофическая функция), поглощают продукты метаболизма, фагоцитируют остатки цитоплазмы формирующихся сперматозоидов (резидуальные тельца) и дегенерирующие половые клетки, секретируют жидкость для транспорта сперматозоидов в семенных канальцах. Сустентоциты (клетка Сертоли) 1.Цилиндрические клетки с гофрированной апикальной и латеральной поверхностями,куда погружены половые клетки на разных стадиях развития 2.Боковые отростки связаны плотными контактами, разделяющими сперматогенный эпителий на базальной и адлюминальное пространства (гематотестикулярный барьер) 3.Физическая поддержка половых клеток 4.Трофическая функция 5.Фагоцитоз клеточных остатков после спермиогенеза 6.В мембрану встроены связанные с G-белком рецепторы ФСГ 7.Секретируют в кровь эстрогены, ингибин, мюллеров ингибирующий фактор 8.Синтез АСБ в просвет канальца, фактор стволовых клеток, трансферрин 9.Секреция жидкости для транспорта сперматозоидов в семенных канальцах. Вопрос 7. Гематотестикулярный барьер. Его структура и функция. Гематотестикулярный барьер. Поддерживающие клетки в период полового созревания вблизи базальной мембраны образуют плотные контакты, формирующие гематотестикулярный барьер. Благодаря такому барьеру, в адлюминальном пространстве создаётся специфическая гормональная среда с высоким уровнем тестостерона. Барьер изолирует созревающие половые клетки от токсических веществ и препятствует развитию аутоиммунного ответа против поверхностных Аг, экспрессирующихся на мембране созревающих сперматозоидов. Вопрос 8. Эндокринная функция яичка. Мужские половые гормоны и синтезирующие их интерстициальные гландулоциты (клетки Лейдига), их строение и локализация, участие в регуляции сперматогенеза. Эндокринная функция (синтез мужских половых гормонов) — в интерстициальных эндокринных клетках. Гипоталамо-гипофизарная система при помощи гонадолиберина активирует синтез и секрецию гонадотропных гормонов гипофиза, влияющих на активность поддерживающих эпителиальных и интерстициальных эндокринных клеток. В свою очередь вырабатываемые в яичке гормоны корректируют эндокринную деятельность гипоталамо-гипофизарной системы. Гонадолиберин Гонадолиберин поступает в кровь из аксонов нейросекреторных клеток в пульсирующем режиме с пиковыми интервалами около двух часов. Гонадотропные гормоны, как и гонадолиберин, высвобождаются в кровь также в пульсирующем режиме, что особенно характерно для лютропина, пики концентрации которого в крови мужчин наблюдаются с интервалами 90–120 мин. Гонадотропные гормоны Секреция гонадотропных гормонов поддерживается гонадолиберином, а угнетается тестикулярными гормонами. Тормозящий эффект тестостерона на секрецию лютропина в основном проявляется на уровне гипоталамуса (через синтез гонадолиберина), тогда как эстрогены снижают чувствительность гонадотропных клеток к гонадолиберину. Половые стероиды слабо влияют на секрецию фоллитропина, в то время как пептидный гормон ингибин оказывает выраженное подавляющее действие на синтез фоллитропина. Мишени гонадотропных гормонов — яички. Сустентоциты имеют рецепторы фоллитропина, а интерстициальные эндокринные клетки — лютропина. Фоллитропин активирует в поддерживающих эпителиальных клетках синтез и секрецию ингибина и эстрогенов. Лютропин стимулирует в интерстициальных эндокринных клетках синтез и секрецию тестостерона. Пролактин. Механизмы участия пролактина в регуляции функций яичка не очень ясны, хотя секреция адекватного количества пролактина кажется необходимой для синтеза тестостерона. Увеличение уровня пролактина в крови ведёт к подавлению синтеза тестостерона. У больных с выраженной гиперпролактинемией в результате снижения секреции тестостерона развивается импотенция. Тестикулярные гормоны Тестостерон поступает в сустентоциты, в цитозоле связывается с АСБ и далее секретируется в адлюминальное пространство, для поддержания сперматогенеза. Часть тестостерона конвертируется в эстрогены. Тестостерон и эстрогены крови оказывают ингибирующий эффект на секрецию лютропина гонадотрофами аденогипофиза. Эстрадиол. В гладком эндоплазматическом ретикулуме сустентоцитов путём ароматизации происходит превращение тестостерона, синтезированного в интерстициальных эндокринных клетках, в эстрогены. Эстрогены связываются с рецепторами в интерстициальных эндокринных клетках и по паракринному механизму подавляют синтез тестостерона. Ингибин. В ответ на стимуляцию фоллитропином клетки сустентоциты секретируют ингибин, блокирующий синтез фоллитропина гонадотропными клетками аденогипофиза. Структура ингибина гомологична MIF, секретируемому фетальными сустентоцитами. Эндокринные интерстициальные клетки (Лейдига) 1.Локализуются между извитыми семенными канальцами 2.Развита эндоплазматическая сеть, многочисленные митохондрии, липидные включения, палочковидные кристаллы Райнке 3.В мембрану встроены связанные с G-белком рецепторы лютропина Тестостерон 1.5α-редуктаза → дигидротестостерон 2.3α -редуктаза → андростендиол 3.Циркулирует в крови в комплексе со стероидсвязывающим глобулином или альбумином 4.В плодном периоде определяет развитие плода по мужскому типу 5.В постнатальном обеспечивает развитие вторичных половых признаков, сперматогенез, функцию простаты, семенных пузырьков, бульбоуретральных желёз, рост мышечной массы, хряща, окостенение эпифизарной пластинки, увеличение содержания ЛПНП и снижение ЛПВП 6.Ароматизация тестостерона в печени, жировой и нервной тканях с помощью фермента Р450 ароматазы приводит к образованию эстрадиола и эстрона 7.Деградация в печени, экскреция с мочой Взаимодействие клетки Лейдига и клетки Сертоли Вопрос 9. Гормональная регуляция сперматогенеза. Вклад гормонов гипоталамуса и аденогипофиза. Гормональная регуляция сперматогенеза Гонадолиберин поступает в кровь из аксонов нейросекреторных клеток в пульсирующем режиме с пиковыми интервалами около двух часов. Гонадотропные гормоны, как и гонадолиберин, высвобождаются в кровь также в пульсирующем режиме, что особенно характерно для лютропина, пики концентрации которого в крови мужчин наблюдаются с интервалами 90–120 мин. • Фоллитропин стимулирует в сустоцитах секрецию андрогенсвязывающего белка (АСБ), а также ингибина, который блокирует секрецию фоллитропина гонадотрофами аденогипофиза по принципу отрицательной обратной связи. • Лютропин активирует в интерстициальных эндокринных клетках секрецию тестостерона. Часть тестостерона поступает в кровоток, а другая часть в сустентоцитах соединяется с АСБ и секретируется в адлюминальное пространство, или путём ароматизации превращается в эстрогены. Циркулирующие в крови тестостерон и эстрадиол оказывают отрицательный обратный эффект на секрецию лютропина. Вопрос 10. Семявыносящие пути. Строение и функции. Семявыносящие пути представлены системой канальцев яичка и его придатка, по которым семенная жидкость продвигается в мочеиспускательный канал и включает в себя прямые канальцы, непосредственно отходящие от извитых и впадающие в канальцы сети яичка, которое располагается в средостении. От сети отходят 12-15 извитых выносящих канальцев, которые образуют головку придатка семенника и там соединяются с единственным протоком придатка, он, многократно извиваясь, формирует тело придатка, и в его хвостовой части переходит в прямой семявыносящий проток, который вблизи мочевого пузыря заканчивается ампулярным расширением. Сразу за ампулой после слияния с семенными пузырьками семявыносящий проток переходит в семявыбрасывающий проток, проходящий через предстательную железу и впадающий в простатическую часть уретры В стенке семявыносящих путей выделяют три оболочки с разной степенью развития в различных отделах путей, слизистую, мышечную и адвентициальную. Эпителий слизистой прямых канальцев сети семенника и извитых канальцев однорядный с признаками железистой деятельность в извитых канальцах группы высоких реснитчатых клеток чередуются с низкими железистыми клетками, секретирующими по апокриновому типу. Благодаря этому просвет извитых выносящих канальцев неровный (фестончатый). В протоке придатка эпителий становится двухрядным, а просвет - ровным. Продвижение семенной жидкости по семявыносящим путям обеспечивается мерцанием ресничек эпителиоцитов и медленными перистальтическими сокращениями гладких миоцитов мышечной оболочки, толщина которой растет по направлению к семявыбрасывающему протоку. Слизистая оболочка семявыносящего протока представлена собственным слоем рыхлой соединительной ткани и выстлана многорядным эпителием. Мышечная оболочка образована тремя слоями гладкомышечной ткани, причем во внутреннем и наружном слое гладкие миоциты расположены продольно, а в среднем слое - циркулярно. Мощная мышечная оболочка семявыносящего протока способствует выбрасыванию сперматозоидов во время эякуляции. Адвентициальная оболочка образована волокнистой соединительной тканью с кровеносными сосудами нервами и группами гладких миоцитов. В составе семенного канатика семявыносящий проток проходит по паховому каналу в брюшную полость. В отличие от семявыносящего протока семявыбрасывающий проток не имеет столь мощно выраженной мышечной оболочки, а его адвентициальная оболочка срастается с соединительно-тканной стромой простаты. Вопрос 11. Придаток яичка. Строение канальцев и функции. В придатке яичка различают головку (сaput epididymidis), тело (сorpus epididymidis) и хвост (сauda epididymidis). Головка придатка представлена 10–12 выносящими канальцами (ductuli efferentes). Тело и хвост придатка образованы протоком придатка (ductus epididymis), в который открываются ductuli efferentes. Выносящие канальцы придатка Выносящие канальцы придатка прободают белочную оболочку (tunica albuginea) и соединяют сеть яичка (rete testis) с протоком придатка яичка. Они выстланы эпителием, клетки которого имеют разную высоту (гирляндный эпителий). Высокие цилиндрические клетки снабжены ресничками, способствующими перемещению сперматозоидов по канальцам. Низкие кубические клетки имеют складчатую поверхность с микроворсинками. В апикальной части клеток присутствуют многочисленные пиноцитозные пузырьки и лизосомы. Функция этих клеток заключается в реабсорбции жидкости, образующейся в извитых канальцах яичка. Снаружи эпителиальной выстилки располагается собственный слой с циркулярно ориентированными ГМК, также способствующих продвижению сперматозоидов по выносящим канальцам. Проток тела придатка Проток тела придатка — одиночный и сильно извитой каналец длиной 4–6 м. Он выстлан многорядным цилиндрическим эпителием В эпителии различают два типа клеток: базальные вставочные и высокие цилиндрические. Цилиндрические клетки снабжены стереоцилиями, склеенными в виде конуса (пламенный эпителий). Между основаниями цилиндрических клеток расположены мелкие вставочные клетки, являющиеся предшественниками цилиндрических клеток. Под эпителием располагается собственный слой, окружённый циркулярно ориентированными ГМК. Мышечный слой протока придатка по мере приближения к ductus deferens становится более выраженным и представлен тремя слоями: внутренним и наружным продольными и средним циркулярным. Сокращения ГМК способствуют продвижению сперматозоидов в семявыносящий проток. 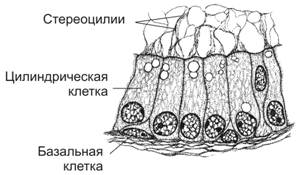 (Кратко) Выносящие канальцы (ductuli efferentes). Выстланы эпителием, клетки которого имеют разную высоту (гирляндный эпителий). Высокие цилиндрические клетки снабжены ресничками, способствующими перемещению сперматозоидов. Низкие кубические клетки имеют микроворсинки и содержат лизосомы. Функция этих клеток заключается в реабсорбции жидкости, образующейся в яичках. Проток придатка (ductus epididymis). 1 Одиночный и сильно извитой каналец длиной 6 метров. Место функционального созревания и запасания сперматозоидов. 2В эпителии различают базальные и цилиндрические клетки со стереоцилиями (пламенный эпителий). 3 Реабсорбция жидкости, фагоцитоз погибших сперматозоидов, секреция сиаловой кислоты и глицерофосфохолина, ингибирующих капацитацию Вопрос 12. Семявыносящий проток. Строение и функции. Из придатка сперматозоиды попадают в семявыносящий проток (ductus deferens). Толстая стенка ductus deferens состоит из трёх оболочек: слизистой, мышечной и адвентициальной. Слизистая оболочка представлена собственным слоем слизистой и многорядным эпителием, который в проксимальной части семявыносящего протока по строению сходен с эпителием протока придатка. Мышечная оболочка образована тремя слоями ГМК: внутренним и наружным продольными и средним циркулярным. Мощная мышечная оболочка ductus deferens способствует выбрасыванию сперматозоидов во время эякуляции. Адвентициальная оболочка состоит из волокнистой соединительной ткани с кровеносными сосудами, нервами и группами ГМК. В составе семенного канатика ductus deferens проходит по паховому каналу в брюшную полость. Вблизи мочевого пузыря семявыносящий проток заканчивается ампулярным расширением. Сразу за ампулой после слияния с семенными пузырьками ductus deferens продолжается в семявыбрасывающий проток. 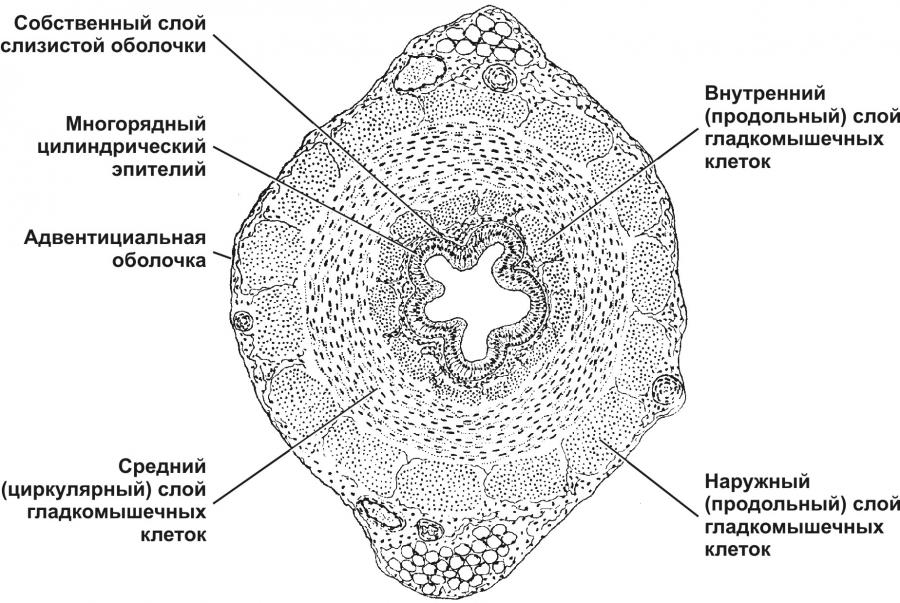 Семявыносящий проток (ductus deferens). Имеет толстую стенку, состоящую из слизистой, мышечной и адвентициальной оболочек. Слизистая оболочка представлена собственным слоем и эпителием, напоминающим эпителий протока придатка. Мышечная оболочка образована внутренним и наружным продольными и средним циркулярным слоями ГМК. Сокращение ГМК способствует выбрасыванию сперматозоидов во время эякуляции Вопрос 13. Семенные пузырьки. Строение и функции. Семенные пузырьки — две извитые трубки длиной до15 см, открывающиеся в семявыбрасывающий проток (сразу за ампулой семявыносящего протока). Стенка трубки состоит из трёх оболочек: слизистой, мышечной и соединительнотканной. Слизистая оболочка имеет выраженную складчатость и выстлана однослойным многорядным цилиндрическим эпителием (секреторные и базальные клетки). Мышечная оболочка образована внутренним циркулярным и наружным продольным слоями ГМК. Секрет: 1. Вязкая желтоватого цвета жидкость (70% эякулята), поступает в семявыбрасывающий проток во время эякуляции. 2. Секрет разжижает семя, содержит аминокислоты, фруктозу, соли аскорбиновой и лимонной кислот, холин, простагландины — т.е. вещества, обеспечивающие сперматозоиды энергетическим запасом, повышающие их выживаемость и функциональную активность. 3. Фруктоза и кристаллы холина маркёры семенной жидкости в судебной медицине. Вопрос 14. Бульбо-уретральные железы. Строение и функции. Бульбоуретральные железы (glandula bulbourethralis) — две трубчато-альвеолярные железы небольшой величины, локализующиеся между скелетными мышцами мочеполовой диафрагмы и открывающиеся в губчатую часть уретры. Секреторный отдел железы выстлан эпителиальными клетками кубической и цилиндрической формы, которые продуцируют слизистый секрет, содержащий галактозу, галактозамин, галактуроновая и сиаловая кислоты. В соединительнотканной строме, разделяющей железу на многочисленные дольки, встречаются как ГМК, так и поперечнополосатые мышечные волокна. Предполагают, что вязкий слизистый секрет, выделяемый в период полового возбуждения, служит для смазки уретры перед эякуляцией. Кратко. Бульбоуретральные железы (Купера) — две трубчато-альвеолярные железы, погружённые в скелетную мышечную ткань мочеполовой диафрагмы. Вязкий слизистый секрет (галактоза, галактозамин, галактуроновая кислота) выделяется в период полового возбуждения, служит для смазки уретры перед эякуляцией. Вопрос 15. Предсталельная железа. Строение и функции. Возрастные изменения. Развитие. Дифференцировка и рост предстательной железы находятся под контролем андрогенов. 5a-редуктаза в эпителиальных клетках простаты катализирует превращение тестостерона в дигидротестостерон, ответственный за пролиферацию и дифференцировку клеток железы. До пубертата рост простаты незначителен. В период полового созревания увеличивающийся уровень тестостерона стимулирует развитие и созревание предстательной железы. Строение  Анатомически в железе различают 5 долей: правая и левая латеральные, правая и левая задние и средняя. Гистологически железа состоит из 30–50 разветвлённых трубчато-альвеолярных желёз, расположенных в трёх зонах. Периферическая (содержит самые крупные и большее количество железы), центральная и периуретральная. Железа снаружи покрыта соединительнотканной капсулой, содержащей ГМК. Соединительнотканные перегородки с хорошо развитой гладкой мускулатурой разделяют предстательную железу на дольки. Поскольку каждая отдельная железа простаты — разветвлённая трубчато-альвеолярная, то на срезе концевые отделы имеют самую различную форму: округлую, овальную, вытянутую, разветвлённую. Нередко у мужчин пожилого возраста в просвете секреторных отделов можно обнаружить конкременты — сферической формы тела диаметром до 250 мм, состоящие из белка и солей кальция. Каждая железа имеет собственный выводной проток, открывающийся в просвет уретры. Выводные протоки выстланы многорядным призматическим эпителием, который в дистальных отделах становится переходным. Секреторные отделы выстланы однослойным кубическим или цилиндрическим эпителием (в зависимости от стадии секреции). В составе эпителия присутствуют базальные, железистые и эндокринные клетки. Железистые клетки имеют выраженный белоксинтезирующий аппарат, в апикальной части клеток присутствуют мелкие светлые секреторные гранулы. Клетки продуцируют секрет, содержащий лимонную кислоту, простато-специфичную кислую фосфатазу, фибринолизин, простато-специфический антиген. Эндокринные клетки редко разбросаны среди других эпителиальных клеток. В секреторных гранулах присутствуют соматостатин, серотонин, бомбезин. Базальные клетки обеспечивают регенерацию железистого эпителия. Секрет Секрет предстательной железы попадает в мочеиспускательный канал за счёт сокращения ГМК, он принимает участие в разжижении семени и способствует его прохождению по мочеиспускательному каналу при эякуляции. Присутствующие в секрете предстательной железы протеолитические ферменты [фибринолизин, простато-специфический антиген (сериновая протеаза)] препятствуют склеиванию сперматозоидов. Кратко: Строение. Состоит из 30–50 разветвлённых трубчато-альвеолярных желёз, разделённых содержащими значительное количество ГМК перегородками соединительной ткани. Каждая железа имеет собственный выводной проток,открывающийся в просвет уретры. Секрет попадает в мочеиспускательный канал за счёт сокращения ГМК железы, принимает участие в разжижении семени и способствует его прохождению по мочеиспускательному каналу при эякуляции. В нормальном секрете предстательной железы обнаружены липиды, фибринолизин, простато-специфичный Аг (сериновая протеаза), кислая фосфатаза. Тема 21: «Женская половая система» Вопрос 1. Развитие женской половой системы. Первичные гоноциты, начальная локализация, пути миграции в зачаток гонады. Половая дифференцировка. Органы женской половой системы развиваются из индифферентных гонадных валиков (преимущественно из коркового их вещества) и из внутренних половых протоков (а именно из мюллерова протока).
Первичные гоноциты появляются в стенке желточного мешка. Оттуда мигрируют, образуя в торако-люмбальном отделе нефротома индифферентные гонадные валики. Гонадные валики начинают дифференцироваться в яичники и тут же одновременно в составе этих же яичников в дифференцировку вступают овогонии. Наружные половые органы дифференцируются из мочеполового синуса, полового бугорка, половых складок и половых валиков. Уроректальная перегородка разделяет клоаку на переднюю часть – мочеполовой синус и на заднюю часть, которая впоследствии образует прямую кишку.
Вопрос 2. Яичник. Развитие. Строение. Особенности строения коркового и мозгового вещества яичника. Первичные половые клетки образуются в стенке желточного мешка и мигрируя, на четвертой неделе эмбриогенеза в торако-люмюальном отделе нефротома формируют индифферентные гонадные валики. В отсутствии Y хромосомы, а именно TDF (определяющий развитие яичек фактор, который является продуктом экспрессии гена SRY, локализованного на Y хромосоме) индифферентные гонады дифференцируются в яичники. Яичники образуются преимущественно из корковой части гонадных валиков, мозговое по большей части дегенерирует. Поверхность яичника покрыта одним слоем кубических эпителиальных клеток, находящихся на толстой соединительнотканной пластинке – белочной оболочке яичника. Паренхима яичника состоит из коркового и мозгового вещества. Мозговое вещество образовано рыхлой соединительной тканью, богатой эластическими волокнами, кровеносными и нервными сплетениями. Корковое вещество содержит: - примордиальные фолликулы - первичные, вторичные фолликулы - зрелые фолликулы - желтые тела - белые тела - атретические фолликулы - интерстициальные клетки Атретические фолликулы – дегенерирующие фолликулы, содержащие погибший овоцит (фолликулы проигравшие в борьбе доминантному фолликулу, который овулировал). Полость отсутствует, характерна сморщенная прозрачная оболочка, окруженная дегенерирующими фолликулярными клетками, между которыми находятся волокнистые структуры. Интерстициальные клетки – клетки паренхимы яичника, синтезирующие и секретирующие андрогены. Вопрос 3. Овогенез. Отличия овогенеза от сперматогенеза. Овогенез (образование яйцеклетки) проходит через три стадии: (1) размножения, (2) роста и (3) созревания (мейоз). Первичные половые клетки мигрируют в зачатки яичников и дифференцируются в овогонии. Овогонии сразу вступают в стадию размножения (митоз). Завершив серию митотических делений, овогонии вступают в стадию роста. В этот период в цитоплазме накапливаются желточные включения. Вслед за стадией роста начинается стадия созревания (мейоз). Первое деление мейоза остается незавершённым: образующиеся овоциты первого порядка в профазе первого деления мейоза вступают в длительный период покоя, продолжающийся до наступления половой̆ зрелости. Образуется примордиальный фолликул. С наступлением половой зрелости и установлением овариально-менструального цикла при овуляции завершается первое деление мейоза и начинается второе деление, останавливающееся в метафазе. При этом образуеся крупный овоцит второго порядка и мелкая абортивная клетка — первое полярное (направительное, или редукционное) тельце. Второе мейтическое деление завершается при оплодотворении; овоцит второго порядка делится с образованием зрелой̆ яйцеклетки и второго полярного тельца. Первое полярное тельце также подвергается второму делению мейоза. Таким образом, в ходе овогенеза из одной̆ овогонии образуется одна полноценная яйцеклетка, под прозрачной̆ оболочкой̆ которой̆ локализуются три полярных тельца. Отличия овогенеза от сперматогенеза: 1. Отсутствует фаза формирования; 2. Фазы размножения и роста протекают во внутриутробном периоде; 3. При созревании образуются неодинаковые клетки; 4. Овогенез завершается вне гонад и только при оплодотворении; 5. Большинство клеток гибнет; 6. Процесс цикличный; 7. Овогенез прекращается после менапаузы; 8. В результате образуется только одна гамета. Вопрос 4. Развитие фолликула, его строение на разных стадиях развития. Овуляция. Гормоны гипоталамуса и гипофиза, их роль в регуляции развития фолликулов и овуляции. В плодном периоде первичные половые клетки дифференцируются в овогонии в развивающихся яичниках. Овогонии вступают в стадию размножения. К семи месяцам внутриутробного развития стадия размножения прекращается. Овоциты вступают в стадию роста. После этого овоциты приобретают оболочку из одного слоя фолликулярных клеток, овогенез останавливается в профазе первого митотического деления фазы созревания – образуется примордиальный фолликул. При наступлении периода полового созревания увеличивается секреция гонадолиберина. В условиях низкого уровня эстрогенов гонадолиберин секретируется в высоких дозах в течение 3 – 5 минут с интервалом в 1 час. В ответ на это в передней доле гипофиза начинает секретироваться ФСГ. Под действием ФСГ 3 – 30 примордиальных фолликулов вступают в стадию большого роста, образуя первичные фолликулы (предантральный). Под действием ФСГ фолликулярные клетки начинают пролиферировать и секретировать фермент ароматазу, который катализирует превращение андрогенов в эстрадиол. Увеличивается синтез эстрогенов фолликулярными клетками. Эстрогены стимулируют увеличение количества рецепторов на мембранах фолликулярных клеток. Вторичные фолликулы (антральный). Характеризуются дальнейшим ростом. Один из фолликулов, опережающий в росте все остальные становится доминантным фолликулом. Вокруг фолликулов образуется theca. В theca различают 2 слоя theca externa (представлен соединительной тканью) и theca interna (представлен тека – лютеиновыми клетками). Клетки theca interna секретируют андрогены, которые, проникая через базальную мембрану, вступают в реакцию ароматизации, катализируемую ароматазой. Продуктами этой реакции являются эстрогены. Еще клетки theca interna имеют рецепторы к ЛГ. ЛГ стимулирует синтез клетками theca interna андрогенов. К этому моменту уровень эстрогенов становится на столько высок, что гипоталамус начинает секретировать гонадолиберин в небольших дозах в течение 3 – 5 мин, но с интервалом в 3 часа. В ответ на это гипофиз приостанавливает секрецию ФСГ и начинает активную секрецию ЛГ. Третичный фолликул (Граафов пузырек). При малых концентрациях ФСГ фолликулярные клетки перестают делиться. Но ФСГ продолжает стимулировать транспорт жидкости в полость фолликула, за счет чего фолликул быстро увеличивается в размерах от 200мкм до 2, 5 см. В фолликуле сливаются отдельные полости между фолликулярными клетками, овоцит вытесняется к стенке фолликула – образуется яйценосный бугорок. Под действием ЛГ происходит овуляция и лютеинизируются фолликулярные и theca - лютеиновые клетки – образуется желтое тело. Овуляция происходит: 1. По учебнику. Через 12 часов после достижения ЛГ пика концентрации; 2. По Валлиулину В. В. В период между максимальными концентрациями ФСГ и ЛГ – этот период примерно 12 часов. (Посмотри в лекции даже график рисовали!) Вопрос 5. Овариальный цикл и его гормональная регуляция. Фолликулярная и лютеиновая стадии цикла. Все завязано на двух гормонах эстрогены (эстрон, эстрадиол, эстриол) и гонадолиберин. Синтез и секреция гонадолиберина зависит от концентрации эстрогена в крови женщины. Если концентрация эстрогенов в крови низкая, то в гипоталамусе секретируется в высоких дозах гонадолиберин в течение 3 – 5 минут с интервалом в 1 час. Этот гонадолиберин, транспортируясь по системе малой воротной вены гипофиза, взаимодействует с клетками гипофиза, которые секретируют только ФСГ (т. к. низкая концентрация эстрогенов!). Если же концетрация эстрогенов в крови женщины высокая, то в ответ на это в гипоталамусе секретируется гонадолиберин в небольших дозах в течение 3 – 5 мин, но с интервалом в 3 часа. Этот гонадолиберин так же идет по кровотоку малой воротной вены гипофиза в переднюю долю гипофиза и взаимодействует там с клетками, которые секретируют ЛГ (т. к. концентрация эстрогенов высокая!). Еще В. В. Валиуллин говорил, что есть еще одни клетки в гипофизе, которые секртетируют и ФСГ и ЛГ - они работают при среднем уровне эстрогенов. Фолликулярная стадия. В начале овариального цикла уровень эстрогенов низкий в ответ на это в гипоталамусе секретируется гонадолиберин с известным интервалом, что влечет за собой секрецию в гипофизе ФСГ. ФСГ стимулирует вступление 3 - 30 примордиальных фолликулов в яичнике в стадию роста. Под действием ФСГ фолликулярные клетки начинают делиться и секртеировать фермент ароматазу, который катализирует превращение андрогенов в эстрадиол. Увеличивается синтез эстрогенов фолликулярными клетками. Еще под действием ФСГ жидкость транспортируется в полость третичного (преовуляторного) фолликула из-за чего он увеличивается в размерах с 200 мкм до 1,5 – 2,5 см. Также ФСГ стимулирует появление на мембране фолликулярных клеток рецепторов к ЛГ. Лютеиновая стадия. К середине овариально - менструального цикла концентрация эстрогенов в крови женщины становится такой, что в гипофизе начинают работать клетки, секретирующие ЛГ, а клетки, секретирующие ФСГ, берут отдых. ЛГ стимулирует синтез андрогенов клетками theca interna (эти андрогены проникают через базальную мембрану к фолликулярным клеткам, где под действием фермента ароматазы превращаются в эстрадиол). ЛГ стимулирует овуляцию, формирование желтого тела (и его существование) из фолликулярных клеток, оставшихся в овулировавшем фолликуле, и секрецию клетками желтого тела прогестерона (т. е. в фолликуле уже преимущественно начинает синтезироваться прогестерон, а не эстроген). Если оплодотворение яйцеклетки не происходит, то уровень эстрогена в крови женщины понижается и в результате достигает такого уровня, что в гипофизе начинают свою работу клетки, секретирующие ФСГ. В отсутствии ЛГ происходит инволюция желтого тела, а под действием ФСГ в стадию роста вступают новая порция фолликулов – начинается новый овариально – менструальный цикл. Вопрос 6. Развитие, строение и функции жёлтого тела. Жёлтое тело менструальное и беременности. Развитие. Секреторные клетки желтого тела развиваются из фолликулярных клеток и из клеток theca interna, оставшихся в фолликуле после овуляции. Это происходит под действием ЛГ. В процессе лютеинизации клетки гипертрофируются, в них увеличивается содержание элементов гладкой эндоплазматической сети, накапливаются жировые включения. Васкуляризация жёлтого тела увеличивает поступление холестерина в клетки гранулёзы и влияние лютропина на стероидогенез. В жизни желтого тела выделяют четыре стадии (по Валлиулину): 1. Стадия васкуляризации и пролиферации. 2. Стадия железистого метаморфоза. 3. Стадия расцвета. 4. Стадия инволюции (образование белого тела). Строение. Внутри желтого тела находится соединительнотканный рубец, окруженный лютеиновыми клетками, между которыми расположены капилляры синусоидного типа. Функции. Желтое тело функционирует в лютеиновую фазу овариального цикла. Под действием ЛГ лютеиновые клетки синтезируют и секретируют прогестерон. Также лютеиновые клетки секретируют эстрогены (но прогестерона больше: прогестерона 2/3, а эстрогенов 1/3). Менструальное жёлтое тело функционирует до завершения цикла (имплантации нет). Уровень прогестерона достигает пика через 8–9 дней после овуляции, что приблизительно соответствует времени имплантации (если происходит оплодотворение яйцеклетки). | ||||||||||||||||||||||||||||||