В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

155 этом участке протонефридия и происходит просачивание в каналец жидкости из окружающей паренхимы. Протонефридии образуются за счет эктодермы. Функция их, по-видимому, заключается не столько в удалении из организма жидких продуктов обмена веществ, сколько в регуляции осмотического давления. Этим объясняется особенно сильное развитие протонефридиев у пресноводных турбеллярий. В их ткани вследствие разницы осмотического давления между клетками тела и внешней средой непрерывно поступает вода, избытки которой и выводятся наружу протонефридиями. У примитивных морских турбеллярий (отр. Acoela) специальных органов выделения еще нет и удаление экскретов осуществляется подвижными клетками — амебоцитами, которые, «нагрузившись» продуктами обмена, выходят наружу через кожные покровы. Половая система ресничных червей гермафродитна и нередко сложно устроена (см. рис. 124, 134, 135). Главное усложнение по сравнению с кишечнополостными — образование специальных выводных протоков, служащих для выведения половых клеток наружу. Для большинства турбеллярий характерно также наличие компактных половых желез, обладающих собственными оболочками. Большое разнообразие в строении половой системы турбеллярий не позволяет дать единую характеристику этой системы. Приведенное ниже описание составлено главным образом на базе организации многоветвистых и трехветвистых (Polycla-dida и Tricladida). Мужская система состоит из многочисленных небольших мешочков, семенников, рассеянных в паренхиме. От семенников отходят семявыно-сящие каналы, которые с каждой стороны тела впадают в один продольный проток — семяпровод. Оба семяпровода, идущие по бокам глотки и позади нее, соединившись, образуют семяизвергательный канал, залегающий внутри мускулистого совокупительного органа (penis), который впадает в особое впячивание на теле — половую клоаку; последняя открывается наружу отверстием, через которое совокупительный орган может высовываться наружу. В половую клоаку впадают и женские половые протоки. Женская половая система содержит или много очень мелких яичников, или их только два, или всего один. Уменьшение числа яичников часто происходит оттого, что большинство их утрачивает способность производить настоящие яйцевые клетки и превращается в так называемые желточники, поставляющие лишь питательные вещества. Можно представить себе, как возникло такое изменение в строении и функционировании яичников в процессе эволюции турбеллярий. По-видимому, часть половых клеток каждого из первоначально многочисленных яичников не развивалась, а шла на питание растущих ооцитов (такие отношения бывают у некоторых примитивных турбеллярий). Впоследствии же совершалась дифференцировка яичников на собственно яичник, производящий ооциты, и желточники, клетки которых — желточные клетки — гомологичны ооцитам, но утратили способность к оплодотворению и развитию. Они набиты большим количеством питательных веществ и входят в состав сложного яйца (см. ниже), где служат для питания развивающегося зародыша. От яичников отходят два канала яйцевода, направляющихся кзади и попутно принимающих тонкие протоки жел-точников. Позади глотки оба яйцевода соединяются в один канал — влагалище, который открывается в половую клоаку. Наконец, нередко в половую клоаку впадает еще небольшой мешочек, называемый копуля-тивной сумкой, в которую при оплодотворении поступает семя другой особи. Каждая оплодотворенная яйцеклетка окружается группой жел- 156 точных клеток и вместе с ними покрывается общей скорлупой. В результате образуются сложные (т. е. состоящие из разнородных клеточных элементов) яйца. Они характерны не только для многих турбеллярий, но и для других плоских червей. Оплодотворение внутреннее, причем совокупительный орган одной особи вводится в половую клоаку другой, функционирующей в данный момент как самка. Детали устройства половой системы у разных групп ресничных червей могут варьировать. Так, далеко не у всех турбеллярий происходит разделение женских половых желез на яичники и желточники. Многочисленные семенники нередко подвергаются олигомеризации, и число их низводится до двух (например, в отряде Rhabdocoela). Особого внимания заслуживает крайне примитивно устроенная половая система бескишечных турбеллярий. Половые клетки их лежат прямо в паренхиме, не образуя оформленной железы (см. рис. 134). Нередко отсутствуют и половые протоки. У некоторых примитивных бескишечных турбеллярий (отр. Acoela), по-видимому, еще нет половых отверстий и яйца выводятся через разрыв стенки тела или через рот, что напоминает соответствующий процесс, наблюдаемый у кишечнополостных. Своеобразен и способ оплодотворения, при котором совокупительный орган одной особи вбуравливается прямо в кожу и паренхиму другой, куда и выпускает сперматозоиды. Последние затем уже самостоятельно доходят до яиц и оплодотворяют их. Необычайное разнообразие устройства половых протоков у турбеллярий убедительно свидетельствует, что такая система образований впервые развилась в пределах данной группы. Половые протоки — новоприобретение турбеллярий, связанное с переходом к ползающему образу жизни. Мы застаем турбеллярий в тот момент их эволюции, когда половая система находится «в поисках» новых наиболее выгодных и удобных форм выведения половых продуктов наружу или введения их в тело партнера во время полового процесса. 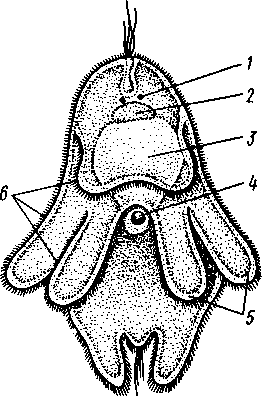 Размножение и развитие. В большинстве случаев размножение только половое. Эмбриональное развитие в разных отрядах ресничных червей сильно различается. Обычно яйца испытывают полное, но неравномерное дробление, напоминающее спиральный тип дробления кольчатых червей (см. далее). Бластомеры яйца еще до образования гаструлы дифференцированы на микромеры, из которых получается в дальнейшем эктодерма, и на 4 макромера, дающих начало энтодерме и мезодерме. У морских турбеллярий отр. Polycladida развитие сопровождается метаморфозом. Из яйца выходит так называемая мюлле-ровская личинка (рис. 132), отличающаяся от взрослой особи многими существенными признаками. Рис. 132. Мюллеровская личинка (из Байера): I — глаза , 2 — мозговой ганглий. 3 — мешковидный кишечник, 4 — ротовое отверстие, 5 — лопасти, 6 — предротовой мерцательный веняик Тело личинки яйцевидно, а не сплющено в спинно-брюшном направлении, кишечник неразветвленный, в виде простого мешка. Особенно же характерно присутствие вокруг середины тела, немного впереди рта, венчика из 8 довольно длинных лопастей, по 157 свободному краю которых проходит пояс сильно развитых ресничек. Совокупность ресничек образует сплошной предротовой мерцательный венчик. Личинка ведет свободноплавающий планктонный образ жизни и, лишь превратившись постепенно в маленькую турбеллярию, опускается на дно. У прочих ресничных червей развитие прямое. 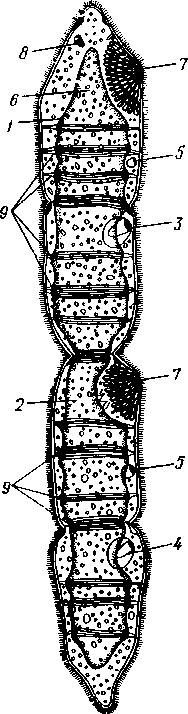 Некоторые турбеллярии (отр. Macrostomida) способны к бесполому размножению посредством повторного поперечного деления. Деление начинается поперечной перетяжкой тела позади глотки (рис. 133). Еще до полного отделения задней половины тела в последней замечается закладка органов, которые впоследствии регенерируют, и образуется целое животное. Позади перетяжки от эктодермы обособляется парная кучка клеток — первый зачаток мозгового ганглия; над ним формируются глаза, а немного позади закладывается зачаток глотки. Только после этого наступает деление. Так как закладка и образование органов происходят заранее, то обе особи (получающиеся в результате деления) сразу же становятся способными питаться, воспринимать раздражения и т. д. У многих турбеллярии отделение задней половины от передней задерживается настолько, что каждый из будущих дочерних индивидов успевает начать подготовку к новому делению. Таким путем у Microstomum и некоторых других составляются цепочки из 4,8 и даже 16 особей, расположенных в один ряд; впоследствии цепочка разрывается на отдельные особи. Такие цепочки носят характер как бы временных колоний. Классификация. До недавнего времени была широко принята старая система классификации турбеллярии, по которой этот класс на основании особенностей строения кишечника делился на отряды бескишечных, многоветвистых, трехветвистых и прямокишечных. В настоящее время классификация основательно переработана и насчитывает 12 отрядов. Назовем лишь главнейшие из них. Рис. 133. Турбеллярия Microstomum (Macrostomida) в состоянии деления — цепочка из 16 особей (из Граф- фа): / — передняя (материн ская) особь, 2 — задняя (1-я дочерняя) особь, 3. 4 — дочерние особи, от делившиеся от двух пер вых и частично уже сформированные, В — начальные стадии отде ления новых дочерних особей, 6 — кишечник, 7 — глотка, 8 — глаза, 9—границы между фор мирующимися особями 158 Отряд 1. Бескишечные (Acoela)—очень мелкие, преимущественно морские формы, характеризующиеся рядом примитивных черт организации (отсутствие кишечника, роль которого выполняет пищеварительная паренхима, отсутствие протонефридиев, поверхностное расположение нервного сплетения, примитивное строение половой системы и др.). Имеют статоцист. Некоторые Acoela содержат в паренхиме многочисленные симбиотические одноклеточные водоросли зоохло-реллы, придающие телу зеленую окраску. Представитель: Convoluta (рис. 134, А),обычная форма морских побережий, объект многочисленных физиологических исследований. Отряд 2. Макростомиды (Macrostomida} — мелкие пресноводные и морские турбеллярии с мешковидным кишечником. Половая система примитивна (нет обособленных желточни-ков). Главные представители: Macrostomum и Microstomum. Последним свойственно бесполое размножение путем поперечного деления, сопровождающегося образованием цепочки особей (см. рис. 133). Отряд 3. Гнатостомулиды (Gnathostomulida)—своеобразная группа мелких турбеллярии, обитающих в песке морских побережий. Имеют мешковидный кишечник и пару хитиноиД-ных челюстей, расположенных в глотке. Характерная особен- ность отряда — строение покровного эпителия, клетки которого несут не реснички, а жгутики (каждая клетка снабжена одним жгутиком). К примитивным чертам строения гнатостомулид следует отнести поверхностное расположение нервной системы, отсутствие протонефридиев, отсутствие обособленных желточников. Представитель: Gnathos-totnula (рис. 134, £). Отряд 4. Многоветвистые (Polycladida) — морские, нередко сравнительно крупные (до 15 см) турбеллярии, обладающие листовидной формой тела. С их крупными размерами связана разветвленное™ кишечника (см. выше) и множественность половых желез (рис. 135). В строении половой системы поликладид есть некоторые примитивные черты — отсутствие обособленных желточников, а у некоторых видов — и половых протоков и т. д. Тропические представители поликладид, особенно обитатели коралловых рифов, характеризуются яркой и пестрой окраской. В наших северных морях обычна Leptoplana (см. рис. 123, В). Развитие идет с метаморфозом (см. рис. 132). Отряд 5. Трехветвистые, или планарии (Tricladida) — многочисленный и богатый представителями отряд. Тело листовидное или лентовидное. Для половой системы характерно наличие многочисленных семенников, двух яичников и множества желточников. Средняя кишка трехветвиста. Преимущественно пресноводные формы, к которым принадлежат наиболее известные наши турбеллярии: молочно-белая планария Dendro-coelum lacteum 2—3 см длины (см. рис. 123, А),темная Polycelis с многочисленными глазами, окаймляющими переднюю половину тела, и др. Немногие Tricladida живут в морях (например, Procerodes lobata с весьма правильным, повторяющимся расположением многих органов по продольной оси тела). Некоторые тропические виды, например Bipalium, приспособлены к наземному образу жизни, встречаются под листьями, в сырой земле и т. п. Наземные Tricladida могут иногда достигать 30 ci/i длины. Особенно богато планариями озеро Байкал, где они достигают значительных размеров и встречаются на глубине до нескольких сотен метров. Отряд 6. Прямокишечные (Rhabdocoela, или Neorhabdocoela). Очень мелкие (0,5— 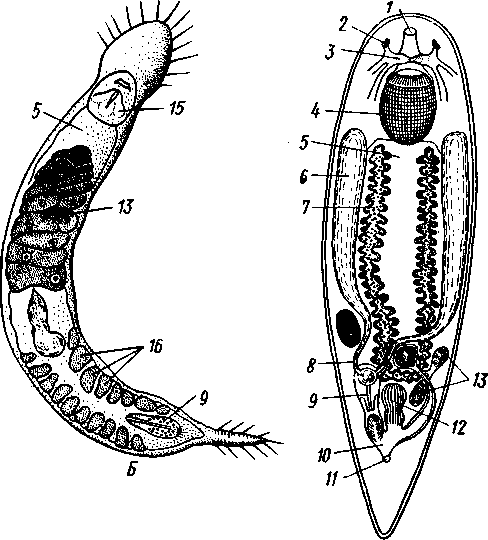 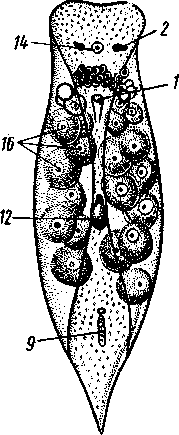 в Рис. 134. Представители разных отрядов турбеллярии. А — бескишечная турбеллярия Convoluta (по Граффу); Б — представитель отряда гнатостомулид Gnatostomula (по Аксу); В — прямокишечная турбеллярия Dalyella viridis (по Бреслау): / — ротовое отверстие, 2 — глаза, 3 — мозговой ганглий, 4 — глотка, 5 — кишечник. 6 — семенники. 7 — желточники, 8 — семяпроводы, 9совокупительный орган, 10 — половая клоака, // — половое отверстие, 12 — копулятивная сумка, 13 — яичники. 14 — статоцист, 15 — хитиноидные челюсти, 16 — половые клетки, лежащие в паренхиме 159 5 мм) животные (см. рис. 134, В) с телом большею частью лишь слабо приплюснутым, почти цилиндрическим или веретеновидным. Многие из них хорошо плавают в воде при помощи биения покрывающих тело ресниц. Характерную черту организации образует прямой, слепо замкнутый сзади кишечник. Ротовое отверстие обычно расположено у переднего конца тела. Над глоткой помещается небольшой мозговой ганглий, от которого идут назад 3, 2 или даже 1 пара 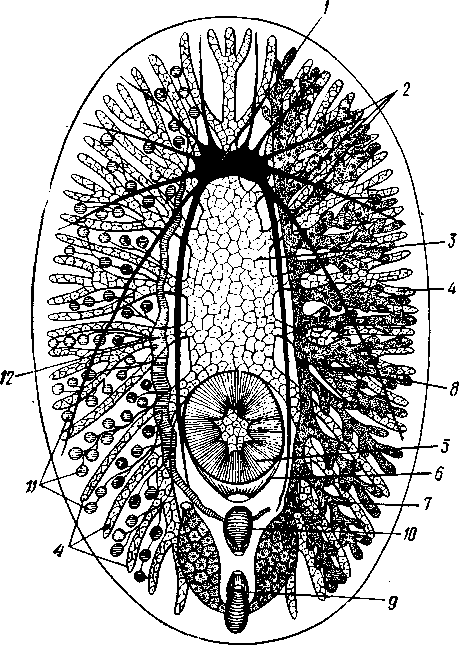 Рис. 135. Организация Polycladida (по Граффу): / — мозговой ганглий, 2 — продольные нервные стволы, 3 — средняя часть кишечника, 4 — боковые ветви кишечника, 5 — глотка. 6 — глоточный карман, 7 — яичники, 8 — яйцевод, 9 — женское половое отверстие, 10 — совокупительный орган. 11 — семенники, 12 — семяпровод. На рисунке слева удалены яичники, справа — семенники нервных стволов. Из органов чувств большей частью имеется пара очень просто устроенных глазков в переднем конце тела. Выделительную систему образует 1 пара каналов с двумя отверстиями. В половой системе характерно массивное строение гонад, которые чаще всего состоят из одной пары семенников, яичников и желточников. Расположение мужского и женского половых отверстий весьма варьирует. При этом в ряде случаев женская половая система помимо протока, служащего для выделения яиц, имеет другой проток специально для совокупления — влагалище. Мы даем столь подробное описание этого отряда ввиду того, что этой группе ресничных червей придается в настоящее время большое филогенетическое значение как исходной группы для других классов червей, с чем мы ознакомимся позже. Прямокишечные встречаются в пресных водах, морях и на суше (во мху). Имеются и паразитические формы, поселяющиеся на моллюсках и других беспозвоночных. Отряд 7. Темноцефалы (Themnocephalida). Эктопаразиты тропических пресноводных ракообразных, моллюсков, водных черепах. Мелкие, несколько миллиметров длиной, уплощенные с вытянутым телом черви, обладающие спереди пучком щупалец (рис. 136, А),а сзади прикрепительной присоской с клейкими железами. Покровы лишены 160 ресничек. В организации темноцефал есть много общего с прямокишечными турбелляриями, тем не менее их систематическое положение спорно. Ранее их помещали среди сосальщиков, позднее выделяли в самостоятельный класс. Сейчас большинство исследователей считают темноцефал турбелляриями, сильно измененными паразитическим образом жизни. Отряд 8. Удонеллиды (Udonellida). Мелкие морские турбеллярии, тело которых имеет лишь несколько миллиметров в длину и несет на заднем конце большую присоску (рис. 136, 5), снабженную клейкими железами (приспособление к паразитическому образу жизни). Удонеллиды поселяются на теле рачков, которые, в свою очередь, являются паразитами рыб. Ротовое отверстие ведет в глотку и кольцевидный кишечник, огибающий комплекс половых желез. Развитие прямое. Систематическое положение удонеллид спорно. 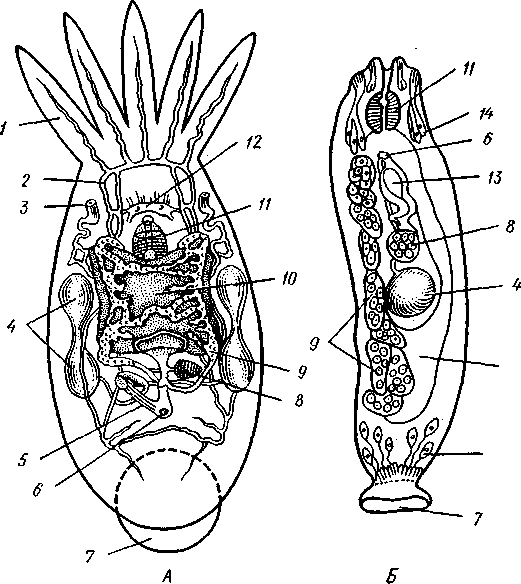 10 /4 яичн мозговой ганг зисты , — 11 глотка, 12 Рис. 136. Паразитические турбеллярии. А — организация Themnocephala (по Бреслау); Б — удонеллиды (Udonellida): 1 — пучок щупалец. 2 — выделительный канал, 3 экскреторное от верстие, 4 — семенники. 5 — копулятивный орган, 6 — половое от верстие, 7 — присоска, 8 — яичник, 9 — желточник, 10 — кишечник, — глий, 13 — зрелое яйцо, 14 — желе ые клетки Происхождение турбеллярии. При рассмотрении турбеллярии мы сталкиваемся с важными вопросами возникновения билатеральной симметрии и происхождения всего типа плоских червей. По вопросу о происхождении турбеллярии наибольшее распространение получили две теории. Долгое время популярностью пользовалась теория Ланга, которая основывается на сходстве организации многоветвистых турбеллярии и гребневиков. И те и другие движутся при помощи ресничек, у обеих групп пищеварительная система устроена по общему типу, состоит из глотки, желудка и отходящих от него слепых ветвей. Подобно гребневикам, некоторые турбеллярии имеют расположенный над мозгом орган 161 6—763 чувства равновесия (статоцист). Наконец, в развитии гребневиков имеются наметки на закладку третьего зародышевого листка — мезодермы, который уже отчетливо выражен у турбеллярий. Сходство турбеллярий и гребневиков казалось очень убедительным при сравнении поликладид с видом Coeloplana metschnikovi, который был описан русским зоологом А. О. Ковалевским. Coeloplana, как и турбеллярий, сплошь покрыта мелкими ресничками, лишена гребных пластинок, листовидно сплющена и может не только плавать, но и ползать по субстрату; кишечник ее сильно разветвлен. По мнению Ланга, эти гребневики обнаруживают известный переход к организации, характерной для отр. Polycladida, и дают ключ к пониманию происхождения плоских червей. Однако теория Ланга во многих отношениях уязвима. В основу филогенетического древа турбеллярий она ставит сложно устроенных поликладид, в то же время примитивных Acoela рассматривает как вторично упрощенных животных. Основываясь на внешнем сходстве и на строении кишечника обеих групп, теория оставляет без внимания особенности строения половой системы и других органов турбеллярий. Кроме того, данные по эмбриональному развитию поликладид и гребневиков, сравнительный анализ соответствия осей тела этих животных показали несостоятельность теории Ланга. Вторая теория происхождения турбеллярий, выдвинутая Граффом и впоследствии модернизированная и дополненная известным советским зоологом В. Н. Беклемишевым, получила более широкое распространение. В этом случае исходным считается отряд Acoela, характеризующийся множеством примитивных признаков организации. Отсутствие кишечника позволяет сравнивать Acoela с планулой (или точнее с паренхимулой) кишечнополостных и высказать предположение о происхождении турбеллярий от каких-то сходных с ней предков. Это могли быть личинки древних кишечнополостных, начавшие размножаться еще до превращения их во взрослую форму (способность к размножению на стадии личинки — явление распространенное и называется неотенией). Дальнейшее усложнение таких неотенических личинок постепенно привело к появлению черт строения, характерных для низших турбеллярий. У паренхимул, как известно, нет кишечной полости, и легко представить себе возможность превращения их энтодермы в пищеварительную паренхиму Acoela. Однако и эта теория, на наш взгляд, не лишена недостатков. Дело в том, что личинка кишечнополостных представляет собой специализированную расселительную личинку, вторично утратившую способность к питанию. Развивающийся из нее полип начинает заглатывать пищу лишь после прикрепления личинки к субстрату и окончательного формирования гастральной полости и ротового отверстия. Приняв теорию Граффа, приходится допустить, что паренхимула не только начала размножаться неотенически, но и вернулась к более примитивному типу питания, который, по-видимому, был свойствен фагоцителлообразным предкам многоклеточных. Эти натяжки отпадают сами собой, если мы примем, что бескишечные турбеллярий ведут свое начало не от неотенических личинок кишечнополостных, а непосредственно от фагоцителлы (с. 93) или подобных ей примитивных многоклеточных организмов. По-видимому, филогенетические взаимоотношения между низшими группами Metazoa (кишечнополостными и бескишечными ресничными червями) можно представить следующим образом. И тип Coelenterata, и примитивные ресничные черви произошли от фагоцителлообразных предков. В первом случае предковые формы прикреплялись к субстрату, теряя при этом способность активно передвигаться. За счет расхожде- 162 ния клеток фагоцитобласта (с. 93) у них формируется гастральная полость, выстланная энтодермой, и прорывается ротовое отверстие. Таким образом, мы приходим к исходному полипоидному организму, характерному для кишечнополостных (с. 116), и только у их расселительной личинки сохраняются отчетливо выраженные признаки предковых форм (с. 577). Во втором случае первичные многоклеточные организмы не прикреплялись к субстрату, а ложились на него боковой поверхностью тела, сохраняя при этом способность к передвижению. Появление постоянного ротового отверстия, превращение фагоцитобласта в пищеварительную паренхиму и развитие примитивных форм полового размножения приводят нас к ацелообразным предкам двусторонне-симметричных животных, которые, по-видимому, и дали начало настоящим бескишечным тур-белляриям. Отсутствие у последних специальных органов выделения, оформленных (одетых оболочкой) гонад и половых протоков, выведение половых продуктов путем разрыва стенки тела или через рот — все эти и другие примитивные особенности Acoela могут найти свое объяснение в исходной простоте организации их фагоцителлообразного предка. Переход первичных форм к придонному ползающему образу жизни привел к изменению лучистой симметрии тела на билатеральную. У прикрепленных и пассивно плавающих планктонных животных все стороны тела вокруг главной оси находятся в одинаковом отношении к захвату пищи, раздражению внешними стимулами и т. д. Когда же подвижные фагоцителлообразные организмы оказались на дне, эти отношения изменились. Сторона тела, соприкасающаяся с субстратом, стала брюшной, а конец, направленный при ползании вперед, оказался в особых условиях в смысле получения раздражений извне, что послужило причиной развития морфологических отличий спинной и брюшной сторон тела, а на переднем конце постепенно сконцентрировались нервные элементы и образовался мозг. Рот, который у исходных форм, так же как и у планул кишечнополостных, прорывался на заднем конце тела, с переходом предков турбеллярий к ползанию, вероятно, сместился на брюшную поверхность. Это должно было облегчить подбирание пищевых частиц с субстрата. Однако дифференцировка переднего конца тела, на котором оказались сосредоточены органы чувств, сделала более выгодным (в смысле поисков пищи, а с переходом к хищничеству — и захвата добычи) переднее положение ротового отверстия. Очевидно, в ходе эволюции турбеллярий происходил процесс постепенного смещения рта, сохранившего свое положение на брюшной стороне, к переднему концу тела. Отражение этого процесса можно видеть в разнообразии положения ротового отверстия у разных групп современных турбеллярий. Совокупностью всех перечисленных изменений в конце концов определилось возникновение у предков ресничных червей билатеральной симметрии. В пределах самого класса турбеллярий эволюция пошла по пути развития нервной системы и органов выделения, совершенствования половой системы и других систем органов. Большую роль в этих процессах играло явление олигомеризации (уменьшалось исходно большое число нервных стволов, выделительных каналов, глаз, гонад и т. д.). |