В.А. Догель - Зоология беспозвоночных. Тип губки (spongia, или porifera)
 Скачать 15 Mb. Скачать 15 Mb.
|

|
РАЗДЕЛ БИЛАТЕРАЛЬНЫЕ (BILATERIA) ТИП ПЛОСКИЕ ЧЕРВИ (PLATHELMINTHES, ИЛИ PLATODES) 1. Тип плоских червей представлен двусторонне-симметричными (билатеральными) животными, через тело которых можно провести только одну плоскость симметрии. Двусторонняя симметрия впервые появляется именно в этой группе беспозвоночных. Плоские черви трехслойны. В процессе онтогенеза у них формируются не два, как у кишечнополостных, а три зародышевых листка. Между эктодермой, образующей покровы, и энтодермой, из которой построен кишечник, у них имеется еще и промежуточный зародышевый листок — мезодерма. Тело их в большинстве случаев вытянуто в длину и сплющено в спинно-брюшном направлении (принимает вид листа, пластинки, ленты). 2. Важная особенность строения плоских червей — наличие у них кожно-мускульного мешка. Так называется совокупность эпителия и расположенной непосредственно под ним сложной системы мышечных волокон. Эти волокна, нередко распадающиеся на несколько слоев (кольцевые, продольные), одевают под эпителием все тело животного в виде сплошного мешка, а не разбиваются на отдельные мускульные пучки более специального назначения, как у высших билатеральных животных (членистоногих, моллюсков). Сокращением мышечных элементов кожно-мускульного мешка обусловливаются характерные «червеобразные» движения Plathelminthes. 3. Тело плоских червей не имеет полости — это бесполостные, или паренхиматозные, животные: пространство между внутренними органами заполнено соединительной тканью мезодермального происхождения, или паренхимой, содержащей многочисленные клетки. Паренхима занимает все промежутки между органами, и ее роль многообразна. Она имеет опорное значение, служит местом накопления запасных питательных веществ, играет важную роль в процессах обмена и т. д. 4. Пищеварительный канал имеет еще примитивное устройство, состоя лишь из эктодермальной передней кишки, или глотки, и энтодер-мальной средней кишки, замкнутой слепо. Задней кишки и заднепроходного отверстия нет. У паразитических форм пищеварительная система может полностью редуцироваться. 5. Нервная система состоит из парного мозгового ганглия и идущих от него кзади нервных стволов, соединенных кольцевыми перемычками. Особого развития достигают два продольных ствола (боковые или брюшные). У плоских червей формируется центральный регулирующий аппарат нервной системы. 6. Кровеносная и дыхательная системы отсутствуют. 7. Впервые появляются специальные органы выделения, построенные по типу так называемых протонефридиев. Они представлены системой 148 разветвленных канальцев, оканчивающихся в паренхиме особой звездчатой клеткой с пучком ресничек. С внешней средой протонефридии сообщаются при помощи специальных экскреторных (выделительных) отверстий. 8. Половая система плоских червей гермафродитна; как правило, формируется сложная система протоков, служащих для выведения половых продуктов, и появляются органы, обеспечивающие возможность внутреннего оплодотворения. К типу плоских червей принадлежат 5 классов: ресничные черви (Turbellaria), сосальщики, или трематоды (Trematoda), моногенеи (Мо-nogenoidea), ленточные черви (Cestoda) и цестодообразные (Cestoda-ria). Последние 4 класса представлены исключительно паразитическими формами. КЛАСС I. РЕСНИЧНЫЕ ЧЕРВИ (TURBELLARIA) Свободноживущие морские или пресноводные, редко наземные, черви, все тело которых покрыто мерцательным эпителием (рис. 123). Число видов турбеллярий достигает 3000. 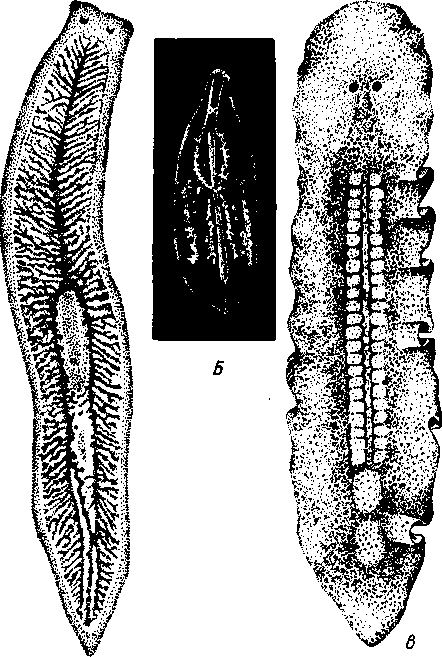 Строение и физиология. Тело вытянуто в длину, листовидно и обыкновенно лишено каких бы то ни было придатков. Лишь у немногих турбеллярий на переднем конце тела имеются небольшие щупальцевидные выросты (рис. 124). Величина турбеллярий незначительна и варьирует от миллиметра до нескольких сантиметров. Тело лишь в редких случаях бесцветно или сплошного белого цвета; обычно животное окрашено в разные, иногда очень яркие цвета зернами залегающего в коже пигмента. Рис. 123. Ресничные черви (Turbellaria). A — Dendrocoelum lacteum (Tricladida); Б — Mesosto- та ehrenbergi (Rhabdocoeia); В — Leptoplana al- cinoi (Polycladida) (по Стрелкову) Тело покрыто однослойным мерцательным эпителием (рис. 125, А), который имеет вид высоких цилиндрических клеток, несущих по свободному краю реснички. У некоторых турбеллярий границы между отдельными клетками исчезают. В таких случаях участки цитоплазмы, несущей ядра, иногда довольно глубоко погружаются в толщу тканей, приобретая вид 149 колбочек, которые суженной частью соединены с наружным слоем (рис. 125,5). Последний лишен ядер и представляет собой неразделенную цитоплазматическую пластинку с ресничками по свободному краю. Такой тип строения покровов называется погруженным эпителием. 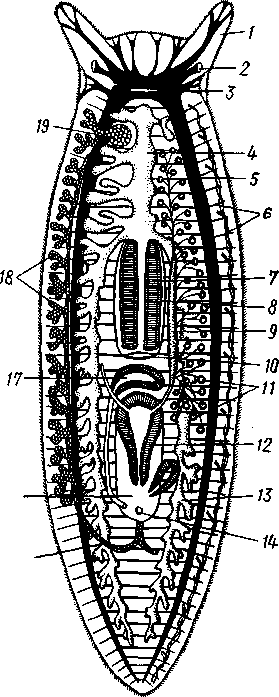 16 15 С покровами турбеллярий связаны многочисленные кожные железы. Обычно это бокаловидной или грушевидной формы железистые клетки, разбросанные по всей поверхности тела или собранные в комплексы. Они могут залегать как в самом эпителии, так и погружаться в глубь тела. Одной из разновидностей кожных желез являются рабдитные клетки, содержащие блестящие, сильно преломляющие свет палочки — раб-диты. Последние лежат перпендикулярно к поверхности тела. При раздражении животного они легко выбрасываются из эпителия наружу и, сильно разбухая в воде, образуют на поверхности тела червя сгустки слизи, возможно, играющей защитную роль. Под эпителием находится эластичная основная, или базальная, мембрана — продукт выделения оснований эпителиальных клеток; она служит для придания телу определенной формы и для прикрепления мышц. Под базальной мембраной помещается сложная мышечная система, состоящая из нескольких слоев гладких мышечных волокон. Наиболее поверхностно располагается слой кольцевых мышц, несколько глубже — продольных и, наконец, диагональных мышечных волокон, идущих в двух направлениях и перекрещивающихся между собой под углом. Совокупность мышц вместе с эпителием образует кожно-мускульный мешок. Кроме того, у всех турбеллярий имеются характерные для них дорзовентральные, или спинно-брюшные, мышцы. Это пучки волокон, идущих от спинной стороны тела к брюшной и прикрепляющихся к базальной мембране обеих плоских сторон тела; таким образом, животное может сплющивать тело в спинно-брюшном направлении. Рис. 124. Схема строения трехвет-вистой" турбеллярий (по Граффу): 1 — щупальцевидные выросты, 2—глаза, 3 — мозговой ганглий, 4 — передняя ветвь кишечника, 5 — продольный нервный ствол, 6 — поперечные нервные перемычки, 7 — глотка, S — глоточный карман, 9 — семяпровод, 10—ротовое отверстие, // — семенники, 12 — совокупительный орган, 13 — задняя ветвь кишечника, 14 — половое отверстие, 15 — яйцевод, 16—половая клоака, 17 — комулятивная сумка, 18 — желточ-ники, 19 — яичник. Слева удалены семенники, справа — желточники и яичник Движение турбеллярий, с одной стороны,— результат биения ресничек наружного эпителия, с другой — обусловливаются сокращением кожно-мускульного мешка. Turbellaria и ползают, и плавают. Мелкие формы главным образом плавают при помощи ресничек. Более крупные представители ползают, вытягивая, сокращая и изгибая тело. Кнутри от кожно-мускульного мешка все промежутки между органами заполнены паренхимой — рыхлым скоплением соединительнотканых клеток, соединяющихся друг с другом своими отростками. 150 Между клетками имеются небольшие пространства с водянистой жидкостью, что позволяет паренхиме выполнять роль посредника в передаче продуктов пищеварения от кишечника к внутренним органам. Одновременно осуществляется и перенос продуктов обмена к выделительной системе. Кроме того, наличие многочисленных фибриллярных образований позволяет рассматривать паренхиму как опорную ткань. Пищеварительная система состоит из передней и средней кишки, замыкающейся слепо. Рот служит не только для проглатывания пищи, 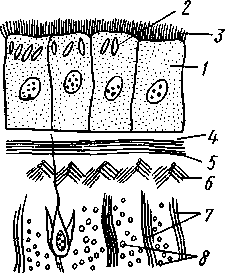 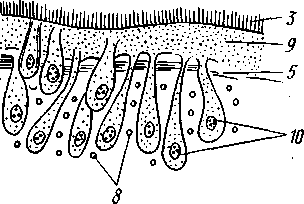 Рис. 125. Схема строения кожно-мускульного мешка. А —с типичным мерцательным эпителием, Б — с погруженным эпителием (по Граф- фу, схематизировано): 1 — клетки эпителия, 2 — рабдиты, 3 — реснички, 4 — базальная мембрана, 5 — кольцевые мышцы, 6 — диагональные мышцы, 7 — дорзовентральные мышцы. 8 — продольные мышцы, 9 — наружный цитоплазматический слой, 10 — погруженные участки цитоплазмы с ядрами  Рис. 126. Строение бескишечной турбеллярии Convoluta (по Бреслау): 1 —• паренхима, 2 — ротовое отверстие, 3 — пищеварительная паренхима, 4 — половые отверстия, 5 — мужской совокупительный орган но и для выбрасывания непереваренных твердых остатков. Большинство турбеллярии — хищники, питающиеся различными мелкими животными. Рот помещается обычно на брюшной стороне тела: на небольшом расстоянии от переднего конца тела, посредине брюшной стороны, или ближе к заднему полюсу. Рот ведет в эктодермальную глотку, которая, в свою очередь, переходит в среднюю кишку. У некоторых крупных турбеллярии (например, у пресноводных планарий отр. Tricladida и у морских турбеллярии отр. Polycladida) ротовое отверстие открывается не в глотку, а в особое глубокое впячивание наружных покровов, называемое глоточным карманом (см. рис. 124, 135). Со дна кармана в его полость вдается мускулистая глотка. Она имеет вид трубки, которая может сильно вытягиваться и высовываться через рот наружу, служа для 151 захвата добычи. Энтодермальная средняя кишка устроена у многих форм Turbellaria различно. У мелких турбеллярий (отр. Rhabdocoela, Macrostomida и др.) она имеет вид простого мешка или слепо замкнутой трубки (см. рис. 134, В).У крупных же форм кишечник, как правило, разветвлен. Так, у многоветвистых (Polycladida) глотка ведет в желудок, от которого во все стороны к краям тела отходят ветвящиеся каналы, слепо замкнутые на концах (см. рис. 135). У трехветвистых (Tricladida) от глотки, находящейся вблизи средины тела, отходят сразу три главные ветви кишечника (см. рис. 123, 124): одна идет прямо вперед, другие две загибаются и по бокам глотки направляются назад; каждая из ветвей дает побочные слепые веточки. Такое строение кишечника не случайно. У мелких турбеллярий, размер которых не превышает нескольких миллиметров, продукты пищеварения легко распределяются по всему телу через рыхлую паренхиматозную ткань. У крупных же три-кладид и поликладид, иногда достигающих 1—3 и даже 30 см, этот процесс чрезвычайно затруднен. У таких турбеллярий функцию распределения продуктов пищеварения по телу животного выполняют ветви кишечника, пронизывающие паренхиму во всех направлениях и непосредственно соприкасающиеся со всеми тканями и органами. В процессе переваривания пищи у турбеллярий, так же как у кишечнополостных, большое место занимает внутриклеточное пищеварение. Частицы пищи, предварительно подвергающиеся обработке секретом глоточных желез, поступают в кишечник и захватываются клетками кишечного эпителия, в которых формируются многочисленные пищеварительные вакуоли. У отряда бескишечных турбеллярий выраженная средняя кишка вообще отсутствует (рис. 126). Глотка у них вдается в несколько обособленный участок паренхимы (пищеварительная паренхима), в которой происходит внутриклеточное переваривание пищи. Нервная система турбеллярий слагается из мозгового ганглия и отходящих от него нервных стволов. Главной особенностью этой системы, по сравнению с таковой кишечнополостных, является концентрация нерв- 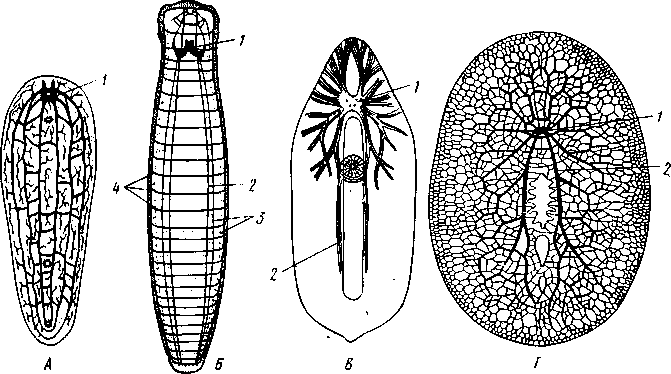 Р'ис. Б :. 127. Нервная система турбеллярий. А — Convoluta (Acocla) (из Байера); — Bothrioplana (представитель отряда, близкого к отр. Tricladida); В — Ме-sostoma (Rhabdocoela); Г — Planocera (Polycladida) (из Беклемишева, по разным _авторам): / — мозговой ганглий, 2 — брюшные продольные стволы, 3 — боковые и спинные продольные нервные стволы, 4 — поперечные перемычки 152 ных элементов у переднего конца тела, сопровождающаяся формированием двойного узла — мозгового ганглия. Это примитивный мозг, выполняющий роль регулирующего нервного центра. От ганглия отходят стволы, состоящие из нервных волокон и рассеянных на их протяжении нервных клеток. Строение нервной системы турбеллярий разнообразно, что позволяет понять основные пути ее эволюции в пределах класса. У некоторых бескишечных турбеллярий (Acoela) нервная система еще очень просто устроена (рис. 127, А).Она представлена диффузным сплетением, расположенным очень поверхностно и несколько напоминающим нервную систему кишечнополостных. Маленький мозговой ганглий иннсрвирует статоцист, расположенный на переднем конце тела червя. Совершенствование этой примитивной нервной системы идет в трех главных направлениях. Прежде всего происходит упорядочивание подкожного сплетения, в результате чего обособляются нервные стволы и соединяющие их поперечные перемычки. Параллельно идет увеличение размеров и усиление роли мозгового ганглия, который из маленького узелка, иннерви-рующего статоцист, постепенно превращается в главный координирующий центр всего тела. Одновременно происходит погружение нервной системы в толщу паренхимы, что можно рассматривать как важное приспособление, обеспечивающее защиту ганглия и стволов от возможных повреждающих воздействий внешней среды. Наконец, следует отметить еще один важный момент в эволюции нервной системы турбеллярий. Число продольных стволов, дифференцирующихся из беспорядочного нервного сплетения, у примитивных форм довольно значительно (5—6 пар). В ходе эволюции оно уменьшается до двух и даже одной пары (рис. 127, В).Оставшиеся стволы обычно сильно развиты и отличаются значительной толщиной. Следовательно, здесь мы имеем дело с явлением олигомеризации гомологичных органов. Все продольные стволы соединены поперечными кольцевыми перемычками, так что нервная система приобретает вид правильной решетки (рис. 127, Б).Такой тип строения нервной системы, свойственный и другим классам плоских червей, называется ортогоном. У своеобразной группы многоветвистых турбеллярий (отр. Polycla-dida) мозговой ганглий сравнительно далеко отодвинут от переднего конца животного и находится между ним и срединой тела (рис. 127, Г), тогда как у трехветвистых (отр. Tricladida), прямокишечных (отр. Rhabdocoela) и некоторых других турбеллярий ганглий приближен к переднему полюсу. Соответственно с этим и расположение нервов, отходящих от ганглия, различно. У многоветвистых турбеллярий они в большом количестве (до 11 пар) расходятся звездообразно во все стороны, причем длина стволов (в связи с положением ганглия) увеличивается от передних к задним. Между соседними нервными стволами имеется система тонких перемычек, образующих сложное нервное сплетение. Два задних нервных ствола развиты более прочих, что придает нервной системе билатеральный характер. У трехветвистых от мозга тоже идет кзади несколько нервных стволов, но среди них особенно выделяется пара брюшных, которые толще всех остальных; таким образом, у трехветвистых билатеральная симметрия в нервной системе сильно выражена. Органы чувств у Turbellaria развиты сравнительно богато. Для осязания служит вся кожа, а у некоторых видов, кроме того, небольшие парные щупальца переднего конца тела. Механические и химические раздражения, поступающие из внешней среды, непосредственно воспринимаются длинными неподвижными ресничками, рассеянными по телу турбеллярий. Эти реснички (или жгутики) устроены принцициально так 153 же, как чувствительный волосок (книдоциль) стрекательных клеток кишечнополостных (с. 118). Однако в отличие от книдоциля они связаны с нервными клетками, отростки которых подходят к мозговому ганглию (рис. 128). Это обеспечивает общую централизованную ответную реакцию животного на раздражение. Органы чувств такого строения назы- 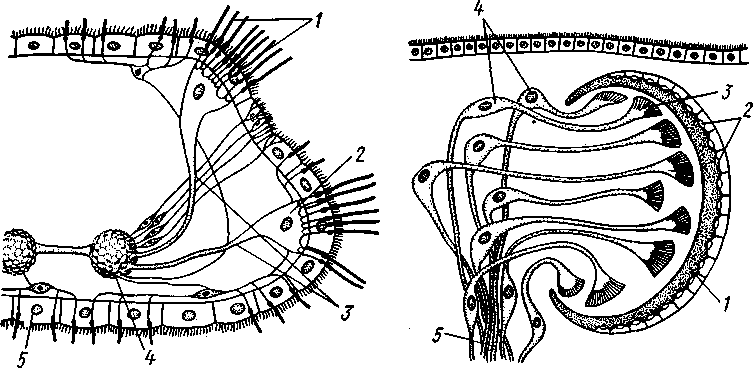 Рис. 128. Схема расположения кожных чувствительных окончаний (сенсилл) тур-беллярии. Поперечный разрез головного конца (схематизировано, по Гелей): 1 — чувствительные реснички, связанные . с нервными клетками, 2 — нервные клетки, 3 — отросток нервной клетки, 4 — мозговой ганглий, 5 — клетка ресничного эпителия Рис. 129. Схема строения глаза турбел-лярии (по Гессе): 7 — пигментный бокал, 2 — ядра клеток пигментного бокала, 3 — светочувствительная часть рецепторных клеток, 4 — ядра рецепторных клеток, 5 — нервные волокна, образующие зрительный нерв ваются сенсиллами. У некоторых турбеллярий (Acoela и др.) имеются органы чувств равновесия в виде замкнутых мешочков — статоцистов со слуховым камешком из углекислой извести внутри. Статоцист помещается над мозговым ганглием. Глаза имеются почти всегда. Глаз может быть одна пара, и они лежат непосредственно над мозгом; или же их больше (несколько десятков), причем они могут окаймлять края всей передней половины тела. Глаз (рис. 129) обычно состоит из пигментного бокала, образованного одной или несколькими пигментными клетками. В полость бокала вдается светочувствительная часть рецепторных клеток. От них отходят нервные волокна, которые соединяются затем в зрительный нерв и направляются к мозгу. Пигментный бокал обращён своей вогнутой стороной к поверхности тела, так что световые лучи должны сначала пройти сквозь тела рецепторных клеток, прежде чем попадут на светочувствительные участки последних. Глаза подобного рода получили название обращенных, или инвертированных. Напомним, что в глазах медуз светочувствительная часть ретинальных клеток, была расположена на тех их концах, которые направлены кнаружи, к источнику света — это глаза необращенные, или неинвертированные. Органов дыхания у турбеллярий нет. Кислород, растворенный в воде, поступает в тело червя путем диффузии через всю поверхность. В этом отношении большое значение имеет уплощенная форма тела турбеллярий. Выделительная система как отдельная система органов впервые появляется у ресничных червей (рис. 130, 131). Она представлена двумя 154 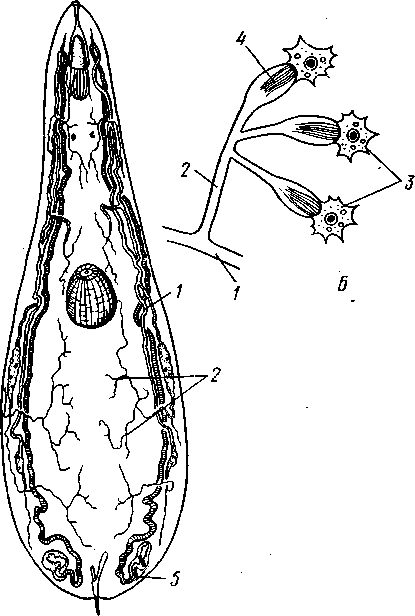 или несколькими каналами, каждый из которых одним концом открывается наружу. От этих главных каналов в глубь тела отходят многочисленные побочные ветви; последние дают начало более тонким протокам. Самые концевые участки всей системы представляют собой очень тонкие канальцы (капилляры), проходящие внутриклеточно (они пронизывают несколько расположенных в ряд клеток). На свободных концах капилляры замыкаются особыми довольно крупными звездчатыми клетками. От внутренней поверхности такой клетки в просвет канальца отходит пучок ресничек. Реснички все время бьют наподобие пламени свечи, колеблемого ветром, почему пучку и дали название мерцатель- 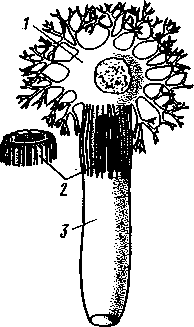 Рис. 131. Электронно-микроскопическая схема строения мерцательной клетки (по Кюммелю, изменено): / — звездчатая часть мерцательной клетки, 2 — цитоплазматические тяжи, 3 — трубка (внутри мерцательное пламя) Рис. 130. Выделительная система турбеллярий. А — расположение главных выделительных каналов в теле прямокишечной турбеллярий Gyratryx hermaphroditus (из Райзингера); Б — схема строения протонефридиев (по Буэ и Шантон): / — главные продольные выделительные каналы, 2 — разветвления канальцев, 3 — звездчатые (мерцательные) клетки, 4 — мерцательное пламя, 5 — выделительные отверстия ного пламени. Постоянная работа ресничек препятствует застою подлежащей удалению жидкости, которая проходит дальше по тонкому выделительному канальцу в силу законов капиллярности. Выделительные органы в виде ветвистых каналов, замкнутых на концах звездчатыми клетками, называются протонефридиями. С помощью электронного микроскопа удалось выяснить ультраструктуру этих образований. Характерная звездчатая часть клетки переходит в трубку (рис. 131), внутри которой и расположен пучок ресничек мерцательного пламени. Начальный участок стенки трубки образован продольно расположенными плотными цитоплазматическими тяжами; тончайшие ще-левидные просветы между ними затянуты слизистой пленкой. Именно в |