ГОРМОНЫ. Учебнометодическое пособие по курсу нормальной физиологии Общие закономерности гормональной регуляции
 Скачать 1.75 Mb. Скачать 1.75 Mb.
|

НОВОСИБИРСКий государственныйМЕДИЦИНСКий университеткАФЕДРА НОРМАЛЬНОЙ ФИЗИОЛОГИИУЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ Общие закономерности гормональной регуляции  НОВОСИБИРСК 2007 г. НОВОСИБИРСКий государственный МЕДИЦИНСКий университет КАФЕДРА НОРМАЛЬНОЙ ФИЗИОЛОГИИ УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ ПО КУРСУ НОРМАЛЬНОЙ ФИЗИОЛОГИИ «Общие закономерности гормональной регуляции» Автор-составитель: профессор кафедры нормальной физиологии НГМУ, д. б. н. Н.Б. Пиковская НОВОСИБИРСК 2007 г. Методическое пособие по курсу нормальной физиологии к разделу: Физиология эндокринной системы «Общие закономерности гормональной регуляции» Новосибирская Государственная медицинская академия, 2007. 76 с. Методическое пособие предназначено для использования в курсе практических занятий по нормальной физиологии студентами второго курса всех факультетов. Утверждено Центральной комиссией методического совета НГМА протокол № от 2007 г. Автор-составитель: профессор кафедры нормальной физиологии НГМА, д. б. н. Н.Б. Пиковская Рецензенты: Зав. кафедрой педагогики и медицинской психологии НГМУ, доцент Г. В. Безродная Профессор кафедры биохимии НГМУ, д .м .н., профессор Н.Н. Маянская
Предисловие Это учебно-методическое пособие составлено в соответствии с лекционным курсом, который в течение последних лет читается на кафедре нормальной физиологии. Прекрасно понимая невозможность в лекционном курсе или коротком пособии изложить все физиологические эффекты гормонов, мы сосредоточили свое внимание на самых общих закономерностях гормональной регуляции, в надежде на то, что эти базовые знания позволят будущим медикам углубить их при необходимости. В своем курсе мы оказались от традиционного изложения свойств отдельных гормонов в соответствии с анатомической принадлежностью, отдав предпочтение общебиологическому значению гормональной регуляции. Поэтому, остановившись коротко на общих закономерностях регуляции секреции гормонов, более подробно рассматриваем значение гормонов в процессах роста и развития организма, регуляции основных параметров гомеостаза (глюкоза, кальций, натрий). Особое внимание и в лекционном курсе, и в пособии уделяется роли гормонов в адаптации, стрессу, как основному механизму адаптации и повреждающим эффектам высоких доз гормонов при длительном стрессе. Изложение физиологии эндокринной регуляции в таком аспекте естественно не позволяет рассмотреть все гормоны организма и все их эффекты, поэтому в качестве приложения в конце пособия дается словарь, в котором представлены физиологические эффекты тех гормонов и биологически активных веществ, которые не обсуждаются в разделах пособия. Введение Две регуляторные системы организма – нервная и гуморальная – выполняя одну функцию в организме человека и животных: приспособление к изменениям во внутренней и внешней среде, - играют свои роли по разному. Если элементарной формой нервной деятельности является рефлекс – мгновенная и точная ответная реакция организма на раздражение рецепторов, которая реализуется путем распространения нервного импульса (потенциала действия), то гуморальная регуляция осуществляется с помощью разнообразных химических веществ, которые с током крови доставляются ко всему организму. Эти вещества называются гормонами. Термин "гормон" был впервые применен в 1902 г. Старлингом и Бейлиссом в отношении открытого ими вещества, продуцирующегося в двенадцатиперстной кишке, - секретина. Термин "гормон" в переводе с греческого означает "побуждающий к действию". Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами, или железами внутренней секреции, и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма. Общие свойства гормонов:
Гормоны выполняют в организме следующие важные функции:
Сравнивая нервную и гуморальную формы регуляции, мы отметили, что ответ гуморальной системы наступает значительно позже, чем рефлекторная реакция. Это связано с тем, что требуется время для секреции гормона, доставки его с током крови к органам мишеням, взаимодействия с мембранными рецепторами. Гуморальная регуляция, в отличие от нервной является не локальной, а генерализованной, потому что рецепторы для гормонов, как правило, находятся в многочисленных тканях и органах. Гуморальная регуляция осуществляется более длительное время, в отличие от кратковременного рефлекторного ответа, и приводит к более выраженным изменениям в тканях, на которые оказывает эффект. В эффектах многих гормонов можно выделить:
Установлены четыре основных типа физиологического действия на организм: кинетическое, или пусковое, вызывающее определенную деятельность исполнительных органов; метаболическое (изменения обмена веществ); морфогенетическое (дифференциация тканей и органов, действие на рост, стимуляция формообразовательного процесса); корригирующее (изменение интенсивности функций органов и тканей). Задумаемся над тем, что изменение метаболизма, или даже структуры органа или ткани приводит к тому, что этот орган, являясь эффектором в рефлекторной дуге, иначе реагирует на возбуждающий импульс. Таким образом, гуморальные механизмы регуляции длительно, в течение часов и суток видоизменяют при необходимости структуры, к которым будут обращены быстрые и точные нервные импульсы. Если учесть, что стимулом для выделения гормонов часто бывает возбуждающий импульс, мы получим представление о взаимодействии нервной и гуморальной регуляции, которые вместе обеспечивают надежное и эффективное приспособление организма к изменяющимся условия как внешней, так и внутренней среды организма. Коротко остановимся на взаимодействии гормонов. Каждый из гормонов или биологически активных веществ выделяется в конкретной ситуации и обладает своим спектром эффектов. Вместе с тем для каждого из гормонов существует постоянный, базовый уровень секреции. В организме человека в норме не бывает такого состояния, когда уровень любого из гормонов равен нулю. Следовательно, гормоны и БАВ могут оказывать друг на друга определенные влияния, и оказывают их. Остановимся на такой форме влияний, которую называют пермиссивные (позволяющие, обусловливающие), или сенсибилизирующие, потенцирующие эффекты. Пермиссивными называются такие эффекты, когда одни гормоны облегчают, или резко усиливают эффекты других гормонов. Например, катехоламины стимулируют гликогенолиз в печени и липолиз в адипоцитах, однако этот эффект не проявляется в отсутствии кортизола. В свою очередь катехоламины потенцируют эффекты глюкокортикоидов. Эстрогены усиливают эффекты многих гормонов, а тиреоидные гормоны увеличивают эффекты эстрогенов. Механизмы таких потенцирующих влияний неспецифичны. Если гормон или БАВ увеличивает проницаемость мембраны клетки для ионов кальция, то такая клетка будет более чувствительна к действию любого вещества. Если гормон повышает проницаемость мембраны для аминокислот и стимулирует синтез белка в клетке, то на мембране клетки увеличивается и количество мембранных рецепторов, специфичных для данной клетки, следовательно, такая клетка становится более чувствительной к действию других гормонов и БАВ. И последнее, на чем мы остановимся во введении: следует понять разницу между метаболическими или клеточными эффектами гормонов и физиологическими эффектами на уровне целого организма. Физиологический эффект – это то, к чему приводят клеточные эффекты гормонов. Например, альдостерон на уровне клеток дистальных канальцев нефрона увеличивает транспорт ионов натрия через клетки путем активации натрий/калиевого насоса, а на уровне организма такой усиленный перенос натрия реализуется в увеличении объема циркулирующей крови, увеличении минутного объема сердца и повышении системного артериального давления. Принципы организации гуморальной регуляции Эндокринная железа - это анатомическое образование, лишенное выводных протоков, единственной или основной функцией которого является внутренняя секреция гормонов. К эндокринным железам относятся гипофиз, эпифиз, щитовидная железа, надпочечники (мозговое и корковое вещество), паращитовидные железы. В отличие от внутренней секреции, внешняя секреция осуществляется экзокринными железами через выводные протоки во внешнюю среду. В некоторых органах одновременно присутствуют оба типа секреции. Инкреторная функция осуществляется эндокринной тканью, т.е. скоплением клеток с инкреторной функцией в органе, обладающем функциями, не связанными с продукцией гормонов. К органам со смешанным типом секреции относятся поджелудочная железа и половые железы. Продолжая сравнение нервной и гуморальной регуляции, вспомним о том, что нервный импульс (ПД) существует столько, сколько продолжается действие раздражителя или реверберация импульса в нервном центре. «Жизнь» гормона в организме не только значительно дольше, но и значительно сложнее. Попробуем пройти вместе с гормоном цикл его существования от синтеза и стимуляции секреции до разрушения и выведения из организма, и выявить наиболее общие закономерности каждого из этапов. Гормональный эффект опосредован следующими основными этапами: синтезом и поступлением в кровь, формами транспорта, клеточными механизмами действия гормонов. Место синтеза и секреции гормонов К эндокринным железам относятся гипофиз, эпифиз, щитовидная железа, надпочечники (мозговое и корковое вещество), паращитовидные железы. В отличие от внутренней секреции, внешняя секреция осуществляется экзокринными железами через выводные протоки во внешнюю среду. В некоторых органах одновременно присутствуют оба типа секреции. Инкреторная функция осуществляется эндокринной тканью, т.е. скоплением клеток с инкреторной функцией в органе, обладающем функциями, не связанными с продукцией гормонов. К органам со смешанным типом секреции относятся поджелудочная железа и половые железы. Одна и та же железа внутренней секреции может продуцировать неодинаковые по своему действию гормоны. Так, например, щитовидная железа продуцирует тироксин и кальцитонин. В то же время продукция одних и тех же гормонов может осуществляться разными эндокринными железами. Например, половые гормоны продуцируются и половыми железами, и надпочечниками; соматостатин секретируется аденогипофизом, поджелудочной железой и клетками кишечника; натриуретический пептид – предсердиями и сосудами, а АДГ только нейрогипофизом. Продукция биологически активных веществ - это функция не только желез внутренней секреции, но и других традиционно неэндокринных органов: почек, желудочно-кишечного тракта, сердца. Не все вещества, образующиеся специфическими клетками этих органов, удовлетворяют классическим критериям понятия "гормоны". Поэтому наряду с термином "гормон" в последнее время используются также понятия гормоноподобные и биологически активные вещества (БАВ), гормоны местного действия. Так, например, некоторые из них синтезируются так близко к своим органам-мишеням, что могут достигать их диффузией, не попадая в кровоток. Такой механизм взаимодействия называют паракринным. Трудность точного определения термина "гормон" особенно хорошо видна на примере катехоламинов - адреналина и норадреналина. Когда рассматривается их выработка в мозговом веществе надпочечников, их обычно называют гормонами, если речь идет об их образовании и выделении симпатическими нервными окончаниями, их называют медиаторами. Регуляторные гипоталамические гормоны - группа нейропептидов, включая недавно открытые энкефалины и эндорфины, действуют не только как гормоны, но и выполняют своеобразную медиаторную функцию. Некоторые из регуляторных гипоталамических пептидов обнаружены не только в нейронах головного мозга, но и в особых клетках других органов, например кишечника: это вещество Р, нейротензин, соматостатин, холецистокинин и др. Клетки, вырабатывающие эти пептиды, образуют диффузную нейроэндокринную систему (в отличие от классической – гландулярной), состоящую из разбросанных по разным органам и тканям клеток. Клетки этой системы характеризуются высоким содержанием аминов, способностью к захвату предшественников аминов и наличием декарбоксилазы аминов. Отсюда название системы по первым буквам английских слов Amine Precursors Uptake and Decarboxylating system - APUD-система - система захвата предшественников аминов и их декарбоксилирования. Поэтому правомерно говорить не только об эндокринных железах, но и об эндокринной системе, которая объединяет все железы, ткани и клетки организма, выделяющие во внутреннюю среду специфические регуляторные вещества. Регуляция секреции гормонов (Механизмы стимуляции или подавления секреции) Говоря о стимулах для секреции гормонов, следует помнить о том, что для всех эндокринных клеток существует постоянный, базовый уровень секреции. Если нам удастся определить концентрацию гормона в крови, то мы убедимся в том, что ни для одного гормона в любой физиологической ситуации не бывает концентраций, равных нулю. Вместе с тем, концентрации гормонов, конечно, изменяются, иначе они не могли бы выполнять своих биологических функций. Что же служит стимулом для изменения темпа секреции уже синтезированного гормона? Таких стимулирующих воздействий несколько. 1. Нервный импульс, например, выделение гипоталамических либеринов и статинов, выделение адреналина и норадреналина мозговым веществом надпочечников резко возрастает при активации симпатической нервной системы. 2. Влияние другого гормона, например, тропные гормоны аденогипофиза (АКТГ, ТТГ, ФСГ и ЛГ) регулируют синтез и секрецию гормонов коры надпочечников. Щитовидной и половых желез, ангиотензин стимулирует все этапы синтеза и секрецитю альдостерона корой надпочечников. 3. Концентрация вещества, уровень которого регулируется гормоном, например концентрация глюкозы в крови служит основным стимулом для выделения инсулина островковыми клетками поджелудочной железы, высокая концентрация ионов кальция в крови непосредственно стимулирует выделение кальцитонина клетками щитовидной железы. 4. Непосредственное раздражение рецепторов (баро-, хемо-), например, стимулом для секреции АДГ в гипоталамусе может быть раздражение гипоталамических осморецепторов, а раздражение механо- и хеморецепторов кишечника приводит к секреции таких гормонов как секретин, гастрин, ХК-ПЗ. 5. Повреждение ткани, например, при травме сосуда вызывает выделение тромбоксана, простациклина; воздействие антигенов стимулирует выделение гистамина, серотонина тканями. Таким образом, для одного гормона возможно несколько вариантов стимуляции и подавления секреции. Такая организация эндокринной регуляции делает ее надежной, и способной обеспечить необходимую перестройку работы организма в любой ситуации. Транспорт Прежде, чем обсуждать варианты транспорта гормонов, вспомним о том, что есть гормоны, которые оказывают свои эффекты непосредственно в том месте, где секретируются, например, гастрин в слизистой желудка. Такое действие называется паракринным. Итак, возможны следующие варианты транспорта гормонов:
Рецепция и клеточные механизмы действия Обязательным условием для проявления эффектов гормона является его взаимодействие с рецепторами. Гормональные рецепторы представляют собой особые белки клетки, для которых характерны: 1) высокое сродство к гормону; 2) высокая избирательность; 3) ограниченная связывающая емкость; 4) специфичность локализации рецепторов в тканях. На одной и той же мембране клетки могут располагаться десятки разных типов рецепторов. Количество функционально активных рецепторов может изменяться при различных состояниях и в патологии. Так, например, при беременности в миометрии исчезают М-холинорецепторы, и возрастает количество окситоциновых рецепторов. Механизмы действия гормонов. Существуют два основных механизма действия гормонов на уровне клетки: реализация эффекта с наружной поверхности клеточной мембраны и реализация эффекта после проникновения гормона внутрь клетки. Механизм действия зависит от биохимической структуры гормона, которая определяет возможность проникновения гормона в клетку. Классификация гормонов и БАВ по химической структуре:
Если гормон не проникает в клетку, он взаимодействует с рецептором, расположенным на поверхности клеточной мембраны. В результате взаимодействия гормона с рецептором активируется система вторичных посредников – специализированных мембранных белков, кооперированных с рецептором, которые изменяют свою конформацию и ферментативную активность при взаимодействии гормона с рецептором (рисунки 1 – 3). Физиологическая роль вторичных посредников заключается в том, что во-первых, многократно увеличивается эффект малых доз гормонов, во-вторых, формируется универсальный ответ клетки при действии различных гормонов. Один из вторичных посредников - мембранный фермент - аденилатциклаза. Этот фермент способствует образованию из аденозинтрифосфорной кислоты (АТФ) важнейшего внутриклеточного посредника реализации гормональных эффектов - циклического 3,5-аденозинмонофосфата (цАМФ). цАМФ активирует клеточный фермент протеинкиназу, реализующую действие гормона изменяя активность различных ферментов путем их фосфориллирования. Установлено, что гормонозависимая аденилатциклаза - это общий фермент, на который действуют различные гормоны, в то время как рецепторы гормонов множественны и специфичны для каждого из них. Вторичными посредниками кроме цАМФ могут быть циклический 3,5-гуанозинмонофосфат (цГМФ), ионы кальция, инозитол-трифосфат. Активация вторичного посредника, как правило, происходит через специальные G-белки, эти белки могут быть как стимулирующими, так и ингибирующими активность вторичного посредника. Так действуют пептидные, белковые гормоны, производные тирозина - катехоламины. Характерной особенностью действия этих гормонов является относительная быстрота возникновения ответной реакции, что обусловлено активацией предшествующих уже синтезированных ферментов и других белков. 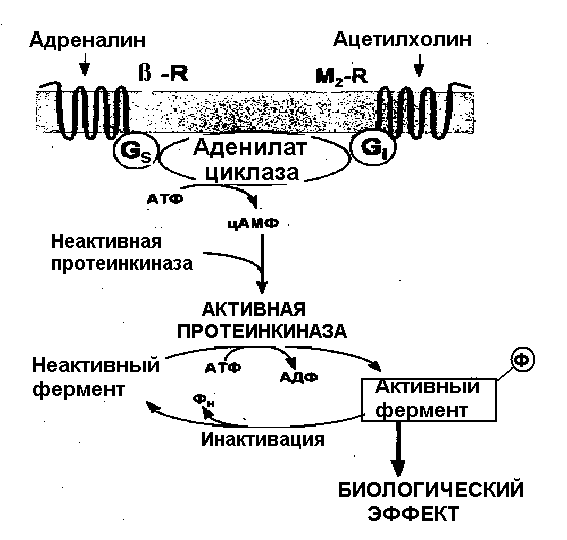 Рисунок 1. Пример стимулирования и ингибирования аденилатциклазы через G-белки двумя различными гуморальными веществами. В случае взаимодействия адреналина со своим рецептором через стимулирующий G-белок происходит активация вторичного посредника – аденилатциклазы, что приводит к активации ферментов и определенному биологическому эффекту. Ацетилхолин, действуя на рецептор через ингибирующий G-белок тормозит этот эффект. 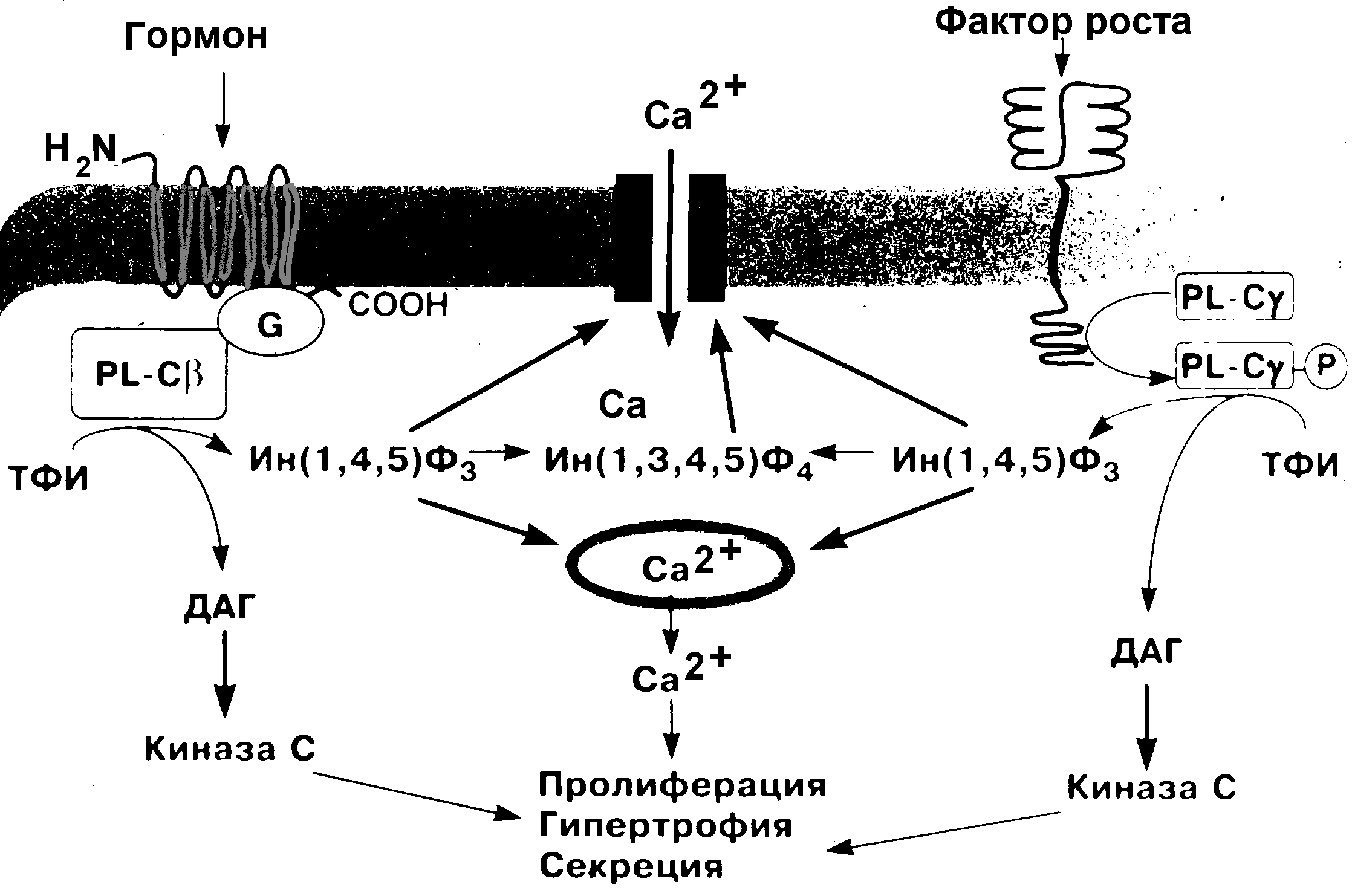 Рисунок 2. Пример (слева) активации гормоном через типичный семидоменный (7 мембранных фрагментов) рецептор G-белка, который активирует фосфолипазу С . Этот универсальный вторичный посредник увеличивает внутриклеточную концентрацию инозитол-трифосфата и в результате концентрацию ионов кальция. Ионы кальция, в свою очередь, активируют ряд метаболических процессов в клетке. Обратим внимание на то, что есть рецепторы (справа), которые обходятся без посредства G-белков, активируют другие фосфолипазы и тоже увеличивает концентрацию в клетке инозитол-трифосфата. Так действуют, например, инсулиноподобные факторы роста через тирозинкиназный рецептор. ТФИ - трифосфоинозитид, ДАГ – диацилглицерол, Ин(1,4,5)Ф3 – трифосфоинозитол. Во втором случае (рисунок 4) рецепторы для гормона находятся в цитоплазме клетки. Гормоны этого механизма действия в силу своей липофильности легко проникают через мембрану внутрь клетки-мишени и связываются в ее цитоплазме специфическими белками-рецепторами. Гормон-рецепторный комплекс входит в клеточное ядро. В ядре комплекс распадается, и гормон взаимодействует с определенными участками ядерной ДНК, следствием чего является образование особой матричной РНК. Матричная РНК выходит из ядра и способствует синтезу на рибосомах белка или белка-фермента. Так действуют стероидные гормоны и производные тирозина - гормоны щитовидной железы. Для их действия характерна глубокая и длительная перестройка клеточного метаболизма. Р 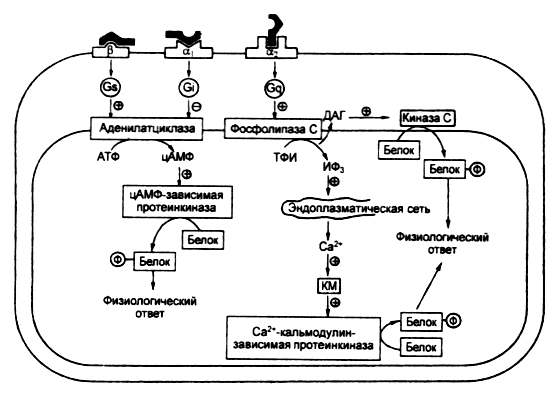 исунок 3. На этом рисунке представлены оба рассмотренных выше варианта на примере взаимодействия адреналина с β- и α-адренорецепторами. Обратите внимание на то, что для реализации эффектов внутриклеточного кальция нужен специальный кальцийсвязывающий белок – кальмодулин, именно этот белок в присутствии кальция и активирует ферменты. 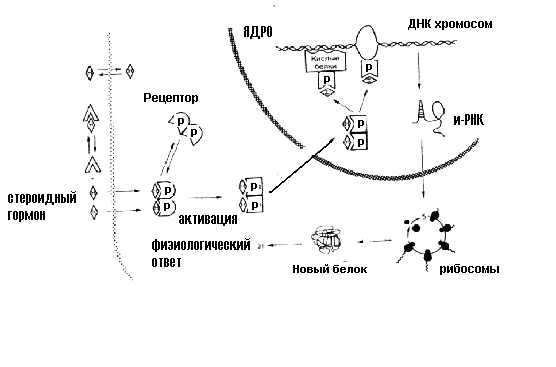 Рисунок 4. Механизм действия стероидных гормонов Для реализации эффекта стероидных гормонов необходим внутриклеточный (а для некоторых гормонов внутриядерный) рецептор, который не только перемещает гормон к хромосомам, но и определяет взаимодействие в определенном месте и экспрессирование генома для синтеза информационной РНК. Физиологические эффекты Следует отличать физиологические эффекты гормонов от механизмов их действия: механизм действия – это то, что происходит в клетках под действием гормонов, физиологический эффект - те изменения, которые при этом происходят на уровне организма. Например, альдостерон увеличивает реабсорбцию натрия в дистальных канальцах нефронов, а на уровне организма задержка натрия приводит к увеличению объема циркулирующей крови и повышению АД. Синтез структурных компонентов клетки под действием соматотропного гормона приводит к росту и развитию всего организма. Метаболизм и выведение Инактивация гормонов происходит в эффекторных органах в результате воздействия ферментов. Большая часть гормонов инактивируется в печени, где гормоны претерпевают различные химические изменения путем связывания с глюкуроновой или серной кислотой. Частично гормоны выделяются с мочой в неизмененном виде. Действие некоторых гормонов может блокироваться благодаря секреции гормонов, обладающих антагонистическим эффектом. Вопросы и задания для самоконтроля
Гипоталамо-гипофизарная система Наиболее сложная и интересная система регуляции секреции гормонов связана с работой гипоталамо-гипофизарной системы. Причин рассмотреть подробно этот вариант регуляции очень много, мы выберем одну, наиболее важную для будущих клиницистов: очень часто нарушения эндокринных функций, приводящие к тяжелейшим клиническим проявлениям, протекают без видимой патологии желез внутренней секреции – нет ни воспалительных процессов, ни травмы, ни опухолей. В то же время изменения концентрации гормонов, их физиологические эффекты, которые в таких случаях становятся уже патологическими, выражены так, что требуют немедленной грамотной коррекции. Дело в том, что в таких случаях речь идет о нарушении регуляции секреции гормонов и изменении механизмов межгоромональных взаимодействий. Гипоталамо-гипофизарная система включает: 1. Клетки гипоталамических ядер, которые обладают секреторной активностью, т.е. могут синтезировать и секретировать в кровь биологически активные вещества. Эти вещества получили название рилизинг-гормонов. Среди рилизинг-гормонов можно выделить либерины – стимуляторы секреции, и статины – ингибиторы. 2. Портальную систему гипофиза – систему капилляров, которая связывает подбугорную область гипоталамуса с аденогипофизом. Именно по этим капиллярам либерины и статины достигают аденогипофиза. 3. Эндокринные клетки аденогипофиза, которые под влиянием рилизинг-гормонов изменяют темп секреции тропных гормонов. Центральная нервная система оказывает свое регулирующее действие на эндокринные железы через гипоталамус. Посредниками между нервной системой и гормональным ответом являются рилизинг-гормоны гипоталамуса. Нейроны гипоталамических ядер обладают уникальной особенностью: их аксоны выделяют свои медиаторы не только к нейронам других областей ЦНС, но и в кровь, через аксовазальные синапсы. Именно выделившиеся в кровь медиаторы гипоталамических нейронов получили название рилизинг-гормонов. Эти гормоны выделяются в капилляры гипофизарной портальной системы и с током крови достигают секреторных клеток аденогипофиза (рисунок 5). 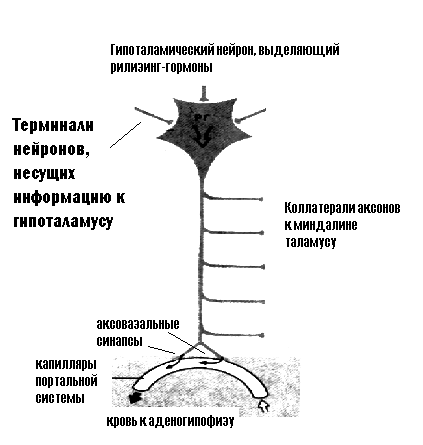 Рисунок 5. Гипоталамический нейрон Каким же образом рилизинг-гормоны включаются в систему гуморальной регуляции деятельности организма? Гипоталамус, который является центром регуляции вегетативных функций, как известно, конвергирует на своих нейронах огромное количество информации. Эти информационные потоки условно можно разделить на несколько групп: а) информация, поступающая по восходящим спинномозговым путям (в основном температурной и болевой чувствительности) от всего организма; б) информация, поступающая по чувствительным веточкам черепно-мозговых нервов – это информация от сердца, сосудов, дыхательной, пищеварительной систем, лица; в) информация, поступающая от органов чувств; г) информация от лимбической системы, которая организует эмоциональную реакцию организма и от коры больших полушарий; д) информация, поступающая не нервным, а гуморальным путем (кровь, мозговая жидкость) о содержании в крови глюкозы, аминокислот, ее осмотической концентрации, температуре, о содержании в крови гормонов. Этот информационный поток обрабатывается ЦНС, приводит к осуществлению определенных безусловных рефлексов, вызывает какие-то изменения поведения и, наряду с этим, стимулирует выделение нейронами гипоталамуса рилизинг-гормонов. Необходимо уточнить, что те из них, которые стимулируют секрецию гормонов клетками гипофиза, называются либерины, а подавляющие секрецию – статины. Рилизинг гормоны, которые с током крови доставляются в аденогипофиз, изменяют темп секреции гормонов аденогипофиза. Эти гормоны, в свою очередь, оказывают влияние на периферические эндокринные железы и ткани организма (рисунок 6).  Рисунок 6. Взаимодействие гипоталамуса и аденогипофиза Таким образом, гипоталамические рилизинг-гормоны представляют собой точку сопряжения во взаимодействии нервной и гуморальной систем регуляции. Такое сопряжение обеспечивает поддержку быстрой и точной, но кратковременной рефлекторной реакции такими изменениями метаболизма, а часто и структуры тканей, которые позволят с максимальной надежностью приспособить организм к изменившимся условиям жизни. Гормоны аденогипофиза можно условно разделить на две группы: гормоны, эффекты которых обращены к периферическим эндокринным железам и регулируют их деятельность и гормоны, которые оказывают влияние непосредственно на ткани организма. К первой группе относятся тропные гормоны: адренокортикотропный гормон (АКТГ) – стимулирует секреторную деятельность коры надпочечников, преимущественно пучковой зоны, тиреотропный гормон (ТТГ) - стимулирует синтез и секрецию гормонов щитовидной железы и гонадотропины, их два – фолликулостимулирующий (ФСГ) и лютеинизирующий (ЛГ) гормоны, которые стимулируют секрецию половых гормонов и регулируют деятельность половой системы. Ко второй группе относятся соматотропный гормон (СТГ) – рост всего организма и пролактин – рост молочных желез. В нейрогипофизе, или задней доле гипофиза гормоны не вырабатываются, туда по аксонам поступают гипоталамические гормоны: антидиуретический гормон и окситоцин, которые из нейрогипофиза выделяются в кровь (рисунок 7). 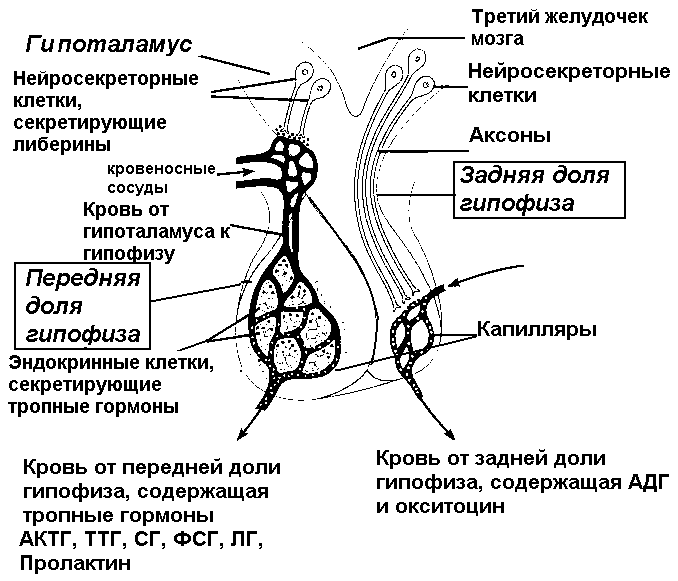 Рисунок 7. Гормоны гипофиза Между рилизинг-гормонами и аденогипофизарными гормонами существует не только система прямых связей, но и обратная связь. Эта связь проявляется в том, что аденогипофизарные гормоны, выделившись в кровь, тормозят секрецию рилизинг-гормонов гипоталамуса. Гормоны аденогипофиза можно объединить в семейства по их химическому строению. Обратите внимание на то, что подобные аденогипофизарным гормоны синтезируются и в плаценте.
Теперь, когда мы знаем названия аденогипофизарных гормонов, будет легко каждому предоставить свой рилизинг-гормон. Либерины и статины гипоталамуса, действующие на аденогипофиз.
Итак, гипоталамические либерины и статины регулируют секреторную деятельность аденогипофиза. Гормоны аденогипофиза оказывают влияние и на железы внутренней секреции, и на ткани организма. В результате повышается секреторная активность желез, находящихся под управлением гипофиза, и метаболизм в тканях. В периферической крови увеличивается уровень гормонов периферических желез и продуктов метаболизма тканей. Эта информация с током крови достигает и гипофиза, и гипоталамуса и приводит к снижению секреторной активности. Таким образом, в гипоталамо-гипофизарной системе работают два варианта обратных связей, в которых информация передается в виде концентрации гормонов: между гипофизом и гипоталамусом короткие обратные связи, а между периферическими железами и гипоталамусом – длинные (рисунок 8). Нейроны гипоталамуса организуют эндокринные функции трех основных гормональных систем:
 Рисунок 8. Прямые и обратные связи в гипоталамо-гипофизарной системе Вопросы и задания
|