Дыхание и обмен веществ. Вопросы коллоквиума по разделам Физиология крови
 Скачать 2.87 Mb. Скачать 2.87 Mb.
|

|
часть кривой демонстрирует относительное постоянство насыщения гемоглобина кислородом, а следовательно, и содержания кислорода в крови независимо от изменений РаО2. Имеется ряд вспомогательных факторов, способствующих оксигенации крови: 1) Отщепление от карбгемоглобина С02 и удаление его (эффект Верига); 2) Понижение температуры в легких; 3) Увеличение pH крови (эффект Бора). 4) Связывание 02 гемоглобином ухудшается с возрастом. Д 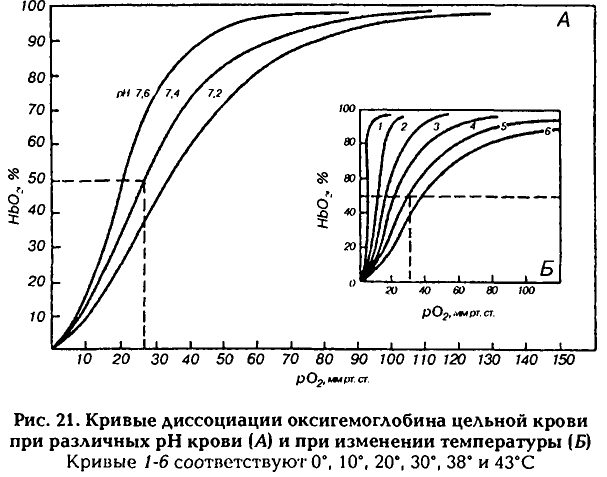 иссоциация оксигемоглобина происходит в капиллярах, когда кровь от легких приходит к тканям организма (это отражает также верхняя часть кривой). При этом гемоглобин не только отдает О2 тканям, но и присоединяет образовавшийся в тканях С02. Главным фактором, обеспечивающим диссоциацию оксигемоглобина, является падение (градиент) Ро2, который быстро потребляется тканями. Процесс диссоциации оксигемоглобина также отражает (как и процесс оксигенации) верхняя часть кривой, так как Р02 в венозной крови составляет 40 мм рт. ст. Эта кровь поступает в легкие, она еще насыщена 02 до 75 %. иссоциация оксигемоглобина происходит в капиллярах, когда кровь от легких приходит к тканям организма (это отражает также верхняя часть кривой). При этом гемоглобин не только отдает О2 тканям, но и присоединяет образовавшийся в тканях С02. Главным фактором, обеспечивающим диссоциацию оксигемоглобина, является падение (градиент) Ро2, который быстро потребляется тканями. Процесс диссоциации оксигемоглобина также отражает (как и процесс оксигенации) верхняя часть кривой, так как Р02 в венозной крови составляет 40 мм рт. ст. Эта кровь поступает в легкие, она еще насыщена 02 до 75 %.В межклеточной жидкости Р02 уменьшается до 5—20 мм рт. ст., а в клетках падает до 1 мм рт. ст. и меньше (когда Р02 в клетке становится равным 0,1 мм рт. ст., клетка погибает). Поскольку возникает большой градиент Р02 (в пришедшей артериальной крови оно около 95 мм рт. ст.), диссоциация оксигемоглобина идет быстро, и 02 переходит из капилляров в ткань. Длительность полудиссоциации равна 0,02 с. Вспомогательные факторы, способствующие диссоциации оксигемоглобина в тканях: 1) накопление С02 в тканях; 2) закисление среды; 3) повышение температуры; 4) АТФ; 5) 2,3-дифосфоглицерат (промежуточный продукт, образующийся в эритроцитах при расщеплении глюкозы, при гипоксии его образуется больше). Миоглобин также присоединяет 02. Сродство миоглобина к 02 больше, чем у гемоглобина: уже при напряжении Ро2 3—4 мм рт. ст. 50% миоглобина насыщено кислородом, а при 40 мм рт. ст. насыщение достигает 95%. Однако миоглобин труднее отдает кислород. Оксимиоглобин начинает отдавать кислород только после того, как Pо2 падает ниже 15 мм рт. ст., благодаря этому он играет в покоящейся мышце роль кислородного депо и отдает 02 только тогда, когда исчерпываются запасы оксигемоглобина, в частности во время сокращения мышцы кровоток в капиллярах может прекращаться в результате их сдавливания, мышцы в этот период используют запасенный во время расслабления кислород. Это особенно важно для сердечной мышцы, источником энергии которой является в основном аэробное окисление. В условиях гипоксии содержание миоглобина возрастает. Сродство к СО у миоглобина меньше, чем у гемоглобина. 24. Транспорт углекислого газа, роль фермента карбоангидразы в транспорте СО2. СО2 транспортируется с помощью плазмы крови и эритроцитов. Основная часть СО2 трансп. плазмой крови, причём около 60% всего Со2 находится в виде бикарбоната NaHCO3 (340 мл/г), 45 мл/г - в виде физ-ки растворённого СО2, 15 мл/г - в виде Н2СО3. Всего в венозной крови 58 мл/г СО2. Несмотря на то, что Рсо2<Ро2, кол-во физически растворённого СО2 в 15 раз больше. Это объясняется более высоким коэф-ом растворимости СО2. В эритроците СО2 находится в форме хим. соединений карбгемоглобина (ННbСО2, 55 мл/г) и бикарбоната КНСО3 (140 мл/г). HHbCO2 обеспечивает выделение из организма до 1/3 объёма всего выделяемого через лёгкие СО2. Это объясняется особенностями строения (Hb-NH-COOH): легко образуется в эритроцитах крови при прохождении её в капиллярах тканей организма и легко диссоциирует, когда кровь проходит в капиллярах лёгки. Практически весь СО2, связанный с гемоглобином, покидает организм с выдыхаемым воздухом в результате газообмена в лёгких. Из др. хим. соед. СО2 освобождается в лёгких лишь частично. СО2, образуемый в организме, выделяется в основном через лёгкие (98%), 0,5% - через почки, около 2% - через кожу в виде НСО3-бкарбонатов. Некоторое увеличение содержания СО2 в крови благоприятно влияет на ораганизм: увеличивается кровоснабжение мозгаи миокарда, стимулируются процессы биосинтеза и регенерации поврежд. тканей, активируется работа сосудодвиг. и дых. центров. Образование соединений СО2 в результате окисл. процессов и образования СО2 его напряжение в клетках и в межклет. простанствах значит. больше (60-80 мм рт. ст.), чем в поступающей к тканям арт. крови (40 мм рт. ст.), поэтому СО2 согласно градиенту напряжения, из интерстиция переходит через стенку капилляров в кровь. Небольшая часть остаётся в плазме в нераствор. виде. В плазме образуется также небольшое количество Н2СО3 (Н20 + + С02 —> Н2С03), но этот процесс идет очень медленно, так как в плазме крови нет фермента карбоангидразы, катализирующего образование Н2С03. Карбоангидраза имеется также в лейкоцитах и тромбоцитах. С02 поступает и в эти клетки, где также образуются угольная кислота и ионы HCO3-. Однако роль этих клеток в транспорте С02 невелика, так как они не содержат гемоглобина, их число значительно меньше, чем эритроцитов, их размеры очень маленькие (тромбоциты имеют диаметр 2 — 3 мкм, эритроциты — 8 мкм). СО2 превращается в др. соединения главным образом в эритроцитах: соедин. с водой и образует угольную кислоту очень быстро (процесс катализируется содержащейся в эритроците карбоангидразой примерно в 15 тыс. раз). Оксигемоглобин, соединенный с ионом калия (КНb02), в тканях с высокой концентрацией С02 легко отдает кислород. В этом состоянии он теряет ионы калия и легко принимает от угольной кислоты ионы водорода, превращаясь в гемоглобиновую кислоту (ННи), в свою очередь ионы калия связываются с ионами гидрокарбоната, образуя бикарбонат калия: КНb02 + Н2СО2 —> ННЬ + КНС03 + 02. Присоединяя ион водорода, гемоглобин действует как буферная система, поэтому большое количество угольной кислоты может переноситься к легким без значительного изменения pH крови. Мембрана эритроцитов слабо проницаема для Na+ и К+, а проникающие ионы транспортируются Na/K помпой обратно: Na+ — в плазму, К+ — в эритроцит. Небольшое количество С02 (1 — 2 %) переносится белками плазмы крови также в виде карбаминовых соединений. Д 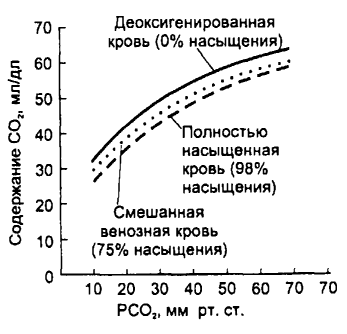 иссоциация соединений С02: в легких происходят обратные процессы, завершающиеся выделением из организма С02 (за сутки выделяется около 850 г С02). В первую очередь начинается выход в альвеолы физически растворенного С02 из плазмы крови, поскольку парциальное давление РС02 в альвеолах (40 мм рт. ст.) ниже, чем в венозной крови (46 мм рт. ст.). Это ведет к уменьшению напряжения РС02 в крови. Присоединение кислорода к гемоглобину также уменьшает сродство углекислого газа к гемоглобину, что облегчает расщепление карбогемоглобина. иссоциация соединений С02: в легких происходят обратные процессы, завершающиеся выделением из организма С02 (за сутки выделяется около 850 г С02). В первую очередь начинается выход в альвеолы физически растворенного С02 из плазмы крови, поскольку парциальное давление РС02 в альвеолах (40 мм рт. ст.) ниже, чем в венозной крови (46 мм рт. ст.). Это ведет к уменьшению напряжения РС02 в крови. Присоединение кислорода к гемоглобину также уменьшает сродство углекислого газа к гемоглобину, что облегчает расщепление карбогемоглобина.Кривая диссоциации С02—гемоглобин значительно отличается от кривой диссоциации оксигемоглобина — она более линейна. Концентрация С02 при любой величине РС02 зависит от степени насыщения гемоглобина кислородом: чем выше насыщение, тем меньше концентрация С02 (эффект Холдейна). (рис) Кривая диссоциации С02 — гемоглобин. Эффект Холдейна. При данном РС02 содержание С02 выше в деоксигенированной крови. 25. Принципы регуляции процесса дыхания (нервный, гуморальный). Понятие дыхательного центра в широком и узком смысле слова (А.А. Ухтомский). Регуляция дыхания – это мероприятия, направленные на сохранение интенсивности потребление О2 и освобождения энергии на относит. постоянном уровне, несмотря на изменение рО2 в окр. среде. Регуляция: 1) рефлекторная; 2) нейрогуморальная. Структура: 1) Рецепторы, воспринимающие инфо. о состоянии системы дыхания и газообмена. 2) Центр. аппарат. 3) Эффекторы (дых. мышцы), неспоредственно осущ. дыхание и связанные каналом обратной связи через рецепторы с центр. аппаратом. Дых. центр – многоуровневая нейронная структура, обеспечивающая регуляцию внеш. дыхания с изменениями во внеш. и внутр. среде. В узком смысле слова дых. центр - совокупность неронов специфических дыхательных ядер продолг. мозга, способных генировать дых. ритм. В широком смысле слова под дых. центром понимают совокупность структур мозга, так или иначе участвующих в регуляции дыхания и в наиболее совершенном приспособлении его к изменяющимся дых. потребностям организма. В физиолог. условиях дых. центр получает афф. сигналы от периф. и центр. хеморецеторов, сигналы о Ро2 в крови и конц. Н+ во внеклеточ. жидкости мозга. В период бодрствования деятельность дых. центра регулируется доп. сигналами, исходящими из различ. структур ЦНС. Нпр., структуры, обеспечивающие речь, т.к. речь может нормы отклонить направление газов в крови от норм. уровня и даже снизить реакци дых. центра на гипоксию или гиперкапнию. Гум. и нерв. принципы регуляции. Афф. сигналы от хеморецепторов тесно взаимодействуют с др. афф. стимулами дых. центра, но в конечном счёте гум. контроль всегда доминирует над нейрогенным. Нпр., человек произвольно не может бесконечно долго задерживать дыхание из-за нарастающих во время остановки дыхания гипоксии и гиперкапнии. Дых. центр выполняет 2 фун-и: 1) Двигательная - регулирует сокращение дых. мышц; закл. в генерации дых. ритма и его паттерна. Генерация дых. ритма - генерация дых. центром вдоха и его прекращение. Паттерное дыхание - длительность вдоха и выхода, величина дых. объёма, минутного объёма дыхания. Моторная фун-я адаптирует дыхание к метабол. потребностям ораганизма, приспосабливает дыхание в поведенч. реакциях, а также осущ. интеграцию дыхания др. фун-ями ЦНС. 2) Гомеостат. фун-я - поддерживает нормальные величины дых. газов (О2, СО2) и рН в крови и внеклет. жидкости мозга, регулирует дыхание при изменении t тела, адаптирует дых. фун-ю к условиям изменёной газовой среды, нпр. при пониженном или повышенном барометрическом давлении. 26. Современные представления о локализации и нейронной организации дыхательного центра. Типы дых.нейронов продолговатого мозга, их классификация. Генерация дыхательного ритма. Классификация нейронов дых. центра: 1) Инспираторные (ранние, возбуждающиеся в начале входа; поздние – в конце; полные – в течене всего вдоха). 2) Экспираторные (ранние, поздние и полные; преинспираторные и постинспираторные). 3) Инспираторно-экспираторные. 4) Экспираторно-инспираторные. 5) Непрерывно активные. Дых. нейроны обнаружены почти на всем протяжении продолг. мозга, однако в обеих половинах продолговатого мозга есть участки РФ, где имеются скопления дых.нейронов. Таких скоплений в каждой половине два — дорсальное и вентральное. Они локализуются вблизи задвижки, которая расположена у нижнего угла ромбовидной ямки. 1) Дорсальная группа дых. нейронов расположена в вентролатеральном отделе ядра солитарного тракта и полностью состоит из инспираторных нейронов — нейронов, возбуждающихся в фазу вдоха, условно - центра вдоха. Аксоны этих нейронов идут к другим нейронам дых. центра и к мотонейронам диафрагм. нерва в передних рогах шейного отдела (C2-4). Нейроны диафрагм. ядра спинного мозга возбуждаются непрерывно, но с учащением в фазу вдоха или залпами, как и связанные с ними нейроны продолг. мозга. Коллатерали от аксонов нейронов дорсального дых. ядра идут также в вентральное дыхательное ядро продолг. мозга, образуя возбуждающие синапсы на его инспираторных нейронах и тормозные — на экспираторных. Экспираторные нейроны в дорсальном ядре встречаются редко — их всего несколько процентов. 2) Вентральная группа дых. нейронов расположена в области обоюдного ядра и простирается от 2-го шейного сегмента сп. мозга включительно, они, подразделяются на две части: - ростральная часть сост. из инспират. нейронов, часть которых взаимодействует с другими дых.нейронами продолг. мозга, другая часть управляет мышцами вдоха с помощью мотонейронов сп.мозга; - каудальная часть состоит из экспираторных нейронов, направляющих аксоны в спинной мозг, мотонейроны которого иннервируют внутренние межреберные мышцы (40 %) и мышцы брюшной стенки (60 %); в вентральной группе находятся также нейроны центров блуждающих нервов. Мотонейроны спинного мозга получают импульсы от центра продолговатого мозга и посылают их к дыхательным мышцам, по диафрагмальному и межрёбер. нервам. Центр диафрагмальных нервов находится в основном в С3-4 спинного мозга. Центры межреберных нервов, иннервирующих мускулатуру грудной клетки локализуются в грудном отделе спинного мозга (Th4-10), иннервация мышц живота осуществляется сегментами Th4-L3. Нейроны моста при взаимодействии с нейронами продолг. мозга и афф. имульсами от лёгких обеспечивают норм. цикл дыхания (эйпностическое дыхания). Отвечает за разговор. Cредний мозг играет важную роль в регуляции тонуса всей мускулатуры организма, в том числе и дыхательной. Гипоталамус выполняет интегрирующую роль в вегеативном обеспечении соматической деятельности, он участвует в регуляции частоты и глубины дыхания при физ. деятельности, повышении температуры внешней и внутр. среды (тепловая одышка). Об участии КБМ в регуляции дыхания свидетельствует тот факт, что частоту и глубину дыхания можно изменять произвольно в широком диапазоне. Но произвольная задержка дыхания не может быть длительной — наступает сильнейшая потребность возобновить его. О роли коры мозга свидетельствует также усиление дыхания перед стартом или по любому условнорефлекторному сигналу. Благодаря КБМ при выполнении физических упражнений интенсивность дыхания становится адекватной потребностям организма (более экономное дыхание). Это связано также и с тем, что сами движения становятся более экономичными. Автоматия дыхательного центра. Под автоматией понимают способность центра к автономной непрерывной генерации импульсов, циркулирующих в его нейронах, обеспечивающих саморегуляцию вдоха и выдоха. Спонтанная активность нейронов дых. центра начинает появл. к концу периода внутриутроб. развития за счёт пейсмекерных св-в сети дых. центра продолг. мозга. Дыхательный цикл подразделяют на фазу вдоха и фазу выдоха. Двум фазам внешнего дыхания соответствуют три фазы активности нейронов дых. центра продолговатого мозга: 1) инспираторная, которая соответствует вдоху; 2) постинспираторная, которая соответствует первой половине выдоха и называется пассивной контролируемой экспирацией; 3) экспираторная, которая соотв. второй половине фазы выдоха и наз. фазой активной экспирации. Инспираторная активность дых. центра начинается с мощного стартового разряда ранних инспираторных нейронов, который появляется спонтанно за 100—200 мс до ]разряда в диафрагмальном нерве. В этот момент ранние инспираторные нейроны полностью освобождаются стосильного торможения со стороны постинспираторных нейронов. Полное растормаживание ранних инспираторных нейронов происходит в период активации преинспираторных нейронов дыхательного центра, окончательно блокирующих разряд экспираторных нейронов. Стартовый разряд ранних инспираторных нейронов активирует полные инспираторные нейроны, которые способны совозбуждать друг друга. Полные инспираторные нейроны благодаря этому свойству поддерживают и увеличивают частоту генерации потенциалов действия в течение фазы вдоха. Именно этот тип дыхательных нейронов создает нарастающую активность в диафрагмальном и межреберных нервах, вызывая увеличение силы сокращения диафрагмы и наружных межреберных мышц. Поздние инспираторные нейроны способны дополнительно активировать в конце вдоха сокращение диафрагмы и наружных межреберных мышц. Прекращение активности всех типов инспираторных нейронов дых. центра растормаживает постинспираторные нейроны. Постинспираторные нейроны регулируют степень расслабления диафрагмы в первую половину фазы выдоха. Вторая половина фазы выдоха, или фаза активной экспирации, полностью зависит от механизма ритмогенеза инспираторной и постинспира-торной активности. Например, при быстрых дыхательных движениях постинспираторная фаза может непосредственно переходить в фазу следующей инспирации. Активность дыхательных мышц. В инспирацию мышечные волокна диафрагмы и наружных межреберных мышц постепенно увеличивают силу сокращения. В этот же период активируются мышцы гортани, которые расширяют голосовую щель. Работа инспираторных мышц во время вдоха создает достаточный запас энергии, которая высвобождается в постинспираторную фазу. В постинспираторную фазу дыхания объем выдыхаемого из легких воздуха контролируется медленным расслаблением диафрагмы и одновременным сокращением мышц гортани. Сужение голосовой щели в постинспиоаторную фазу увеличивает сопротивление воздушному потоку на выдохе. Во вторую фазу выдоха, или фазу активной экспирации, экспираторный поток воздуха усиливается за счет сокращения внутренних межреберных мышц и мышц брюшной стенки. 27. Рефлекторная регуляция дыхания, влияние высших отделов головного мозга на дых. центр. Рефлеторная регуляция дыхания осущ. бюлагодаря обширным связям нейронов дых. центра с многочисл. механорецепторами дых. путей и альвеол лёгких и рецепторов сосудистых рефлексогенных зон. |