Лекции по общей физиологии. 2курс_лекции_по_общей_физиологии. Введение в общую физиологию
 Скачать 6.53 Mb. Скачать 6.53 Mb.
|

|
Тема: Физиология дыхания План: Понятие о процессах дыхания. Механизм вдоха и выдоха. Функциональные показатели дыхания. Обмен газов дыхательной системы. Оксигемоглобин и его диссоциация. Артериально – венозная разность крови по кислороду. Регуляция дыхания. Для нормального протекания жизнедеятельности организма человека необходимо постоянное поступление из внешней среди кислорода и непрерывное удаление из организма углекислого газа, образующегося и накапливающегося в тканях в процессе мета-болизма, потому что в организме человека запасы кислорода ограничены и углекислый газ является токсичным соединением. Сущность дыхания заключается в том, что дыхание является сложным непрерывным процессом, в результате которого посто-янно обновляется газовый состав крови и происходит биологи-ческое окисление в тканях. Нормальное функционирование организма человека возмож-но только при условии пополнения энергией, которая непрерывно расходуется. Организм получает энергию за счет окисления орга-нических веществ – белков, жиров, углеводов. При этом освобож-дается скрытая химическая энергия, которая является источником жизнедеятельности, развития и роста организма. Таким образом, значение дыхания состоит в поддержании в организме опти-мального уровня окислительно-восстановительных процессов. Дыхание состоит из ряда последовательных процессов: внешнее дыхание или легочное дыхание. обмен газов между кровью и легочным воздухом. транспорт газов кровью. обмен газов в тканях организма, потребление кислорода. выделение углекислого газа в клетках в процессе клеточного метаболизма (внутреннее дыхание или клеточное дыхание). Внешнее дыхание – это газообмен между организмом и окружающим его атмосферным воздухом. Внешнее дыхание может быть разделено на два этапа – обмен газов между атмосферным и альвеолярным воздухом и газообмен между кровью легочных капилляров и альвеолярным воздухом. Внешнее дыхание осуществляется за счет активности аппарата внешнего дыхания. Органы внешнего дыхания включает в себя дыхательные пути, легкие, плевру, скелет грудной клетки и ее мышцы, а также диафрагму. Основной функцией аппарата внешнего дыхания является обеспечение организма кислородом и освобождение его от избытка углекислого газа. О функциональном состоянии органов внешнего дыхания можно судить по ритму, глубине, частоте дыхания, по величине легочных объемов, по показателям поглощения кислорода и выделения углекислого газа и т.д. Транспорт газов осуществляется кровью. Он обеспечивается разностью парциального давления газов по пути их следования: кислорода от легких к тканям, углекислого газа от клеток к легким. Внутренне дыхание также может быть разделено на два этапа. Первый этап – это обмен газов между кровью и тканями. Второй связан с потреблением кислорода клетками и выделением ими углекислого газа (клеточное дыхание). Дыхательная система представляет собой совокупность органов выполняющих воздухопроводящую и газообменную функции. В воздухоносных путях происходит увлажнение, согревание, очищение воздуха, газообмен не происходит. Легкие имеют губчатое строение и образованы бронхами, альвеолами и кровеносными сосудами (рис. 5). Альвеола – тонкостенный пузырек, наполненный воздухом и сплетенный сетью капилляров (рис.6). Газообмен происходит через тонкую перегородку, отделяю-щую кровь от альвеолярного воздуха, образованную двумя слоями клеток: стенки капилляра и стенки альвеолы. Общее число альвеол у человека достигает 30 млн. Суммарная площадь поверхности составляет 100 м2, что обеспечивает большую поверхность, через которую происходит диффузия газов в кровь (кислород) и из крови  Рис. 5. Схема воздухоносных путей 1 – гортань; 2- трахея (дыхательное горло); 3 – бронхи; 4 – бронхиальное дерево; 5 - легкое 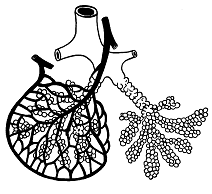 Рис. 6. Схема доли легкого Левая доля оплетена сетью капилляров в альвеолярный воздух (углекислый газ). В дыхательных путях воздух близко не соприкасается с кровью, поэтому газообмен здесь не меняется. Пространство заключенное в этих дыхательных путях называется мертвым пространством. При спокойном дыхании объем воздуха в мертвом пространстве составляет 140-150 мл. Процесс дыхание является ритмичным и цикличным, состоя-щим из актов: вдоха, выдоха и дыхательной паузы. Обычно вдох короче выдоха. Длительность вдоха у взрослого человека 0,9-4,7 с, длительность выдоха 1,2-6 с. Продолжительность вдоха и выдоха зависит в основном от рефлекторных воздействий, идущих от рецепторов легочной ткани. Дыхательная пауза – это непостоянная составная часть дыхательного цикла. Она различна по продол-жительности и даже может отсутствовать. Дыхательные движения совершаются с определенным рит-мом и частотой, которые устанавливаются по количеству экскурсии грудной клетки в 1 мин. У взрослого человека частота дыхательных движений составляет 12-18 в 1 мин. У детей дыхание поверхностное и поэтому более частое, чем у взрослых. У новорожденных детей частота дыхания в покое 40-60 раз в 1 мин. В любом возрасте частота дыхательных движений меньше количества сердечных сокращений в 4-5 раз. При состоянии часто дыхательных движений больше чем при сидении или лежании. Мышечная работа эмоции, повышение окружающей температуры и пищеварение учащают и углубляют дыхание. Во время сна дыхания более редкое. Глубина дыхания падает с увеличением его частоты. На частоту и глубину дыхания влияет обмен веществ. Существует два типа дыхания: грудной (за счет сокращения дыхательной мускулатуры грудной клетки) и брюшной (за счет сокращения диафрагмы). Грудной тип – обладает у женщин, брюшной – у мужчин. Акт вдоха (инспирация) совершается вследствие увеличения объема грудной клетки в трех направлениях – вертикальном, сагиттальном и фронтальном. Изменение размеров грудной полости происходит за счет сокращения дыхательных мышц – наружных межреберных, межхрящных и диафрагмы (рис. 7). 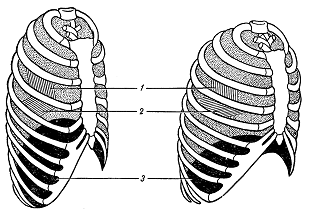 Рис. 7. Схема положения грудной клетки и диафрагмы при выдохе (слева) и вдохе (справа). 1 – наружные межреберные мышцы; 2 – внутренние межреберные мышцы; 3 – диафрагма При сокращении наружных межреберных и межхрящевых мышц, ребра принимают более горизонтальные положения, подни-маясь к верху, при этом нижний конец грудины отходит вперед. Благодаря движению ребер при вдохе размеры грудной клетки увеличивается в поперечном продольном направлении. В результате сокращения мышечных волокон диафрагмы купол ее уплощается и опускается: органы брюшной полости оттесняются вниз, в стороны и вперед, в итоге объем грудной клетки увеличивается в поперечном и продольном направлении. В результате сокращения мышечных волокон диафрагмы купол её уплощается и опускается: органы брюшной полости оттесняются вниз, в стороны и вперед, в итоге объем грудной клетки увели-чивается в вертикальном направлении. При вдохе легкие пассивно следуют за увеличивающейся в размерах грудной клеткой. Дыхательная поверхность легких увели-чивается, давление же в них понижается и становится на 2 мм.рт.ст. ниже атмосферного. Это способствуют поступлению воздуха через воздухоносные пути в легкие Акт выдоха (экспирация) осуществляется в результате расслабления наружных межреберных мышц и поднятия купола диафрагмы. При этом грудная клетка возвращается в исходное положение и дыхательная поверхность легких уменьшается. В начале фазы выдоха давление в легких становится на 3-4 мм рт.ст. выше атмосферного, что облегчает выдох воздуха из них в окружающую среду. Дыхание становится невозможным, когда при ранениях грудной клетки или при разрыве висцерального листка плевры давление внутри легких и вне легких, т.е. между плевральными листками выравнивается давление вследствия поступления воздуха в образующуюся межплевральную щель. Проникновение воздуха между листками плевры называется пневмотораксом, различают односторонний и двусторонний пнев-моторакс при двустороннем полном пневмотораксе, если проколы в грудной клетке широки и легкие спадаются, дыхание прекращается и наступает смерть от удушения. При одностороннем пневмотораксе, если быстро закрыта отверстие в грудной клетке воздух в плевральной полости расса-сывается и дыхание восстанавливается. Человек дышит атмосферным воздухом, который составит 20,9% кислорода, 0,03% углекислого газа, 79,03% азота. В выды-хаемом воздухе обнаруживается 16,3% кислорода, 4% углекислого газа, 79,7% азота. Состав выдыхаемого воздуха весьма непостоянен и зависит от интенсивности обмена веществ, а также от частоты и глубины дыхания. Стоит задержать дыхание или сделать несколько глубоких дыхательных движений, как состав выдыхаемого воздуха изме-няется. Сравнение состава вдыхаемого и выдыхаемого воздуха служит доказательством существования внешнего дыхания. Альвеолярный воздух по составу отличается от атмосфер-ного. В альвеолах происходит обмен газов между воздухом и кровью, при этом в кровь диффундирует кислород, а из крови – углекислый газ. В результате в альвеолярном воздухе резко уменьшается содержание кислорода и возрастает количество углекислого газа. Процентное содержание отдельных газов в альвеолярном воздухе следующее: 14,2-14,6% кислорода, 5,2-5,7% углекислого газа, 79,4-80% азота. Альвеолярный воздух отличается по составу от выдыхаемого воздуха, это объясняется тем, что выдыхаемый воздух содержит смесь газов из альвеол и вредного пространства. Переход газов из окружающей среды в жидкость и из жидкости в окружающую среду подчиняется определенным физическим закономерностям. Каждый газ переходит в жидкость в зависимости от величины его парциального давления. Под парциальным давлением понимают ту часть давления, которая приходит на данный газ в смеси газов. При расчете парциального давления газов в альвеолярном воздухе учитывают его насыщенность водяными парами, парциальное давление которых составляет 47 мм рт. ст. В результате на долю остальных газов альвеолярного воздуха приходится 713 мм рт.ст. Зная процентное содержание газов в альвеолярном воздухе, можно рассчитать их парциальное давление. Для кислорода оно будет составлять 102 мм рт. ст., для углекислого газа 40 мм рт. ст. Движение газов из окружающей среды в жидкость и из жидкости в окружающую среду осуществляется из-за разности их парциального давления. Газ всегда диффундирует из среды, где имеется высокое давление, в среду с меньшим давлением. Это происходит до тех пор, пока не установится динамическое равновесие газов. Самое высокое парциальное давление кислорода в атмосферном воздухе 158 мм рт.ст., в альвеолярном воздухе 108-110 мм рт. ст., а в венозном крови притекающей к легким 40 мм рт. ст. В артериальной крови капилляров большого круга кровообращения напряжение кислорода составляет 102-104 мм рт. ст., в межтканевой жидкости – 40 мм рт.ст., в тканях – 20 мм рт.ст. и меньше в зависимости от функциональной активности клеток. Таким образом, на всех этапах движения кислорода имеется разность его парциального давления, что способствует диффузии газа. Движение углекислого газа происходит в противоположном направлении. Самое большое напряжение углекислого газа имеется в тканях, в местах его образования – 60 мм рт.ст. и более, в венозной крови 46 мм рт.ст., следовательно, разность парци-ального давления углекислого газа по пути его следования является причиной диффузии газа от тканей в окружающую среду. Схема диффузии газов через стенку альвеол представлена на рис. 8. 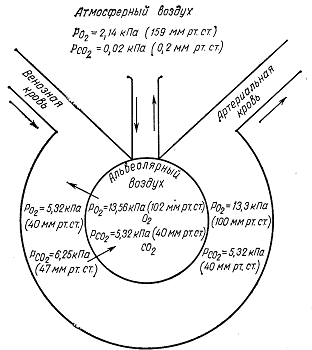 Рис. 8. Схема диффузии газов через мембрану альвеолы В живом организме равенство парциального давления кисло-рода и углекислого газа на этапах их движения никогда не наступает. В легких постоянно происходит обмен газов вследствие дыхательных движений грудной клетки, в тканях же разность парциального давления газов поддерживается непрерывным про-цессом окисления. Проницаемость легочной мембраны для газа выражает величиной диффузионной способности легких. Это – количество газа, проникающего через легочную мембрану за 1 мин на 1 мм рт. ст. градиента давлений. В норме диффузионная способность легких для кислорода около 25 мл/мин. мм рт. ст. Для двуокиси углерода диффузионная способность в 24 раза выше. Диффузионная способность легких зависит: От площади альвеолярно-капиллярной поверхности доступной для газообмена. От толщины легочной и эритроцитарной мембраны. От объема крови в легочных капиллярах и содержание в ни гемоглобина. От пола и возраста человека (у женщин меньше). От положения тела человека в пространстве. В положении стоя диффузионная способность легких на 15-20% меньше, чем в положении сидя, это объясняется перерас-пределением крови в легочных капиллярах. В положении стоя или сидя уменьшается объем крови в верхних долях легких по сравнению с положением лежа, что и приводит к уменьшению диффузионной способности легких. Диффузионная способность легких возрастает при выпол-нении работы. При выполнении работы она может увеличится в 3-4 раза по сравнению с уровнем покоя. Транспорт газов кровью В крови растворяется лишь малая часть кислорода 0,3 мл на каждые 100 мл крови. Основная масса кислорода (98%) находится в связанном состоянии в непрочном соединении с гемоглобином. Всего из крови можно выделить 20 мл кислорода на 100 мл крови. По своему химическому строению гемоглобин относится к группе дыхательных пигментов, в состав которых входят железо-содержащие белки. В нормальных условиях 1 г гемоглобине способен присоединить 1,34 мл кислорода. Средняя концентрация гемоглобина в крови составляет у мужчин – 15 г%, у женщин – 13,5 г%. У спортсменов концентрация гемоглобина увеличивается до 15,5-16,3 г% Одним из основных функциональных показателей крови является кислородная емкость, то есть максимальное количество кислорода, которое могут связать 100 мл артериальной крови при полном насыщении гемоглобина кислородом, которая выражается в объемных процентах (об.%). Средняя величина кислородной емкости у человека равна 19-22 объемных процентов. Количество связанного гемоглобина кислородом более чем в 60 раз превышает количество кислорода, который находится в крови в простом физическом растворе. Степень насыщения гемоглобина кислородом зависит от парциального давления кислорода. При малых величинах напря-жения кислорода происходит расщепление оксигемоглобина, при увеличение напряжения кислорода ускоряется образование окси-гемоглобина. На поглощение О2 гемоглобином влияет также концентрация водородных ионов в крови. Чем больше концентрация водородных ионов, тем меньше сродство гемоглобина к О2. Концентрация водородных ионов зависит главным образом от содержания углекислоты. Чем меньше давление углекислоты в крови, тем больше количество О2 связывается гемоглобином. Наибольшая концентрация водородных ионов в тканях, и меньше в альвеолах. На диссоциацию оксигемоглобина влияет температура. Повы-шение температуры ускоряет только диссоциацию оксигемо-глобина, но практически не влияет на скорость связывания О2 гемоглобином. В тканях при усиленной деятельности органов температура крови несколько повышается, что способствует увеличению отдачи О2 кровью. Перенос О2 гемоглобином в несколько раз выше в физиологических условиях, чем в растворе чистого гемоглобина. Артериальная кровь содержит около 20% об.% (объемных процентов) О2. Проходя через капилляры большого круга крово-обращения, кровь отдает часть содержащегося в ней О2. В покое в венозной крови, оттекающей от тканей, содержится примерно 15 об.%. О2. При физической нагрузке уменьшается до 4-5 об.%. Разность между концентрацией О2 в артериальной и венозной крови называется артерио-венозной разностью крови по кислороду (АВР – О2). Величина АВР – О2 дает представление о количестве О2, используемого тканями. Транспорт углекислого газа. В отличие от кислорода, который связан только с гемогло-бином, для углекислого газа нет одного специального переносчика, он связывает с разнообразными веществами. 1% углекислоты крови составляет свободную углекислоту, 3% ее физически растворено в плазме, около 47 объемных процен-тов связано в плазме в виде углекислых солей (бикарбонатов), в эритроцитах – в виде соединения с гемоглобином и бикарбонатов и некоторое количество углекислоты связано щелочными проте-инами. Углекислый газ в отличие от кислорода может почти беспре-дельно поглощаться кровью, так как в крови есть много оснований для его связывания и превращения в нейтральные соли. В связывании углекислоты главная роль принадлежит белкам крови. Еще И.М. Сеченов установил, что гемоглобин не только косвенно, но и прямо участвует в переносе углекислоты. В тканях образуется непрочное соединение гемоглобина с углекислотой, которая связывается с алеинной группой белка – карбогемоглобин. В присутствии оксигемоглобина карбогемоглобин распада-ется на гемоглобин и углекислый газ. Так в легких выделяется 15-20% всего углекислого газа. Переход оксигемоглобина в гемоглобин в тканях способ-ствует поглощению кровью углекислоты, а переход гемоглобина в оксигемоглобин – выделению углекислого газа из крови в полости легких. В капиллярах малого круга кровообращение под действием карбоангидразы угольная кислота распадается на воду и углекис-лый газ, который диффундирует в альвеолярный воздух. Карбогемоглобин диссоциирует, освобождается СО2, гемогло-бин присоединяет О2, этот процесс называется артериализацией крови. Нервная регуляция дыхательных движений. Дыхательные движения осуществляются благодаря коорди-нированной, согласованной деятельности дыхательных мышц. Нейтроны, дающие начало двигательным нервам дыхатель-ных мышц, расположены в передних рогах спинного мозга. Межреберные нервы исходят из грудных сегментов спинного мозга. Но если отделить спинной мозг от продолговатого мозга, то дыхательные движения прекращаются. Однако они сохраняются, если разрез сделать выше продолговатого мозга. Это доказывает, что согласованная деятельность нейронов двигательных нервов, производящих координированные сокращения дыхательной муску-латуры, вызывается участком нейронов – дыхательным центром, который расположен в продолговатом мозге. Из дыхательного центра нервные импульсы направляются в спинной мозг к центрам двигательных нервов дыхательных мышц и обеспечивают координацию дыхательных движений. Эти нервные импульсы проходят в спинном мозге по передним и переднее – боковым столбам. Н.А. Мисловский (1885) выяснил, что нейроны дыхательного центра находятся главным образом в средней трети продолговатого мозга внутри от ядра подъязычного нерва. В дыхательном центре имеется нейроны, вызывающие вдох (инспираторные) и вызывающие выдох (экспираторные). Возбуж-дение центра вдоха тормозит центр выдоха, и наоборот. Более возбудимы инспираторные нейроны. На поверхности продолго-ватого мозга у основания четвертого желудочка расположен центр, регулирующий частоту дыхания. В варолиевом мосте находятся 2 центра регулирующие дыха-ние: в верхней части мозга расположен «Центр пневмотоксиса», необходимый для нормального дыхания, а в средней и нижней частях – «апнеистический центр», вызывающий судорожные редкие вдохи. После перерезки мозгового ствола ниже этого центра дыхание становится редким и глубоким. Координированные дыхательные движения вызываются толь-ко дыхательным центром продолговатого мозга, который подчиняет себе центры спинного мозга. Дыхание регулируется: нервно-гуморально благодаря возбуждению нейронов дыхательного центра продолговатого мозга колебаниями хими-ческого состава притекающей к ним крови; рефлекторно благодаря притоку афферентных импульсов в дыхательный центр продолговатого мозга. В безусловно-рефлекторной регуляции дыхания участвуют механорецепторы легочной ткани, афферентная импульсация в них возникает при растяжений легочной ткани и возбуждает центр выдоха (рефлекс Геринга-Брейера). Такое влияние оказывает афферентная импульсация с про-приорецепторов межреберных мышц, диафрагма, способствуя чередованию фаз дыхания и обеспечивая соответствие амплитуды дыхательных движений необходимому уровню легочной венти-ляции. Условнорефлекторная регуляция дыхания осуществляется при участии коры больших полушарий, что особенно важно в спортивной деятельности, при адаптации к мышечным нагрузкам. Проявляется в учащении дыхания перед стартом, когда еще нет изменений химизма крови, а также в таких сложных поведенческих реакциях, как речь, пение и др. При нервной регуляции дыхания. Вентиляция легких осуществляется благодаря сокращению дыхательных мышц при возбуждении мотонейронов, которые получают импульсы от инспираторного и экспираторного центров. Их работа регулируется пневматическим центром, находящимся под влиянием гипоталамуса и коры больших полушарий. При гуморальной регуляции дыхания. Изменение состава крови вызывают возбуждение централь-ных и периферических хеморецепторов, афферентная импульсация которых ведет к возбуждению центра вдоха. Периферические хеморецепторы находятся в рефлексогенных зонах дуги аорты и каротидных синусов сонных артерий. Центральные хеморецепторы обнаружены на поверхности продолговатого мозга, стимулируются при гиперкапнии (повыше-нии напряжения СО2 в крови). Центральные хеморецепторы оказы-вают более сильное влияние на деятельность дыхательного центра, чем периферические, усиливают возбуждение центров вдоха и выдоха. Регуляция дыхания при мышечной работе. При усиленной мышечной работе в крови, протекающей через мышцы, увеличивается давление СО2, повышается концентрация водородных ионов и температура крови, что благоприятствует отдаче О2 в мышцах. Эти изменения осуществляются рефлекторно и нервно-гуморально. В результате значительного усиления обмена веществ, поступления в кровь углекислоты, молочной и других кислот и следовательно накопления в крови водородных ионов и возбуж-дения дыхательного центра легочная вентиляция при мышечной работе увеличивается как за счет учащения дыхания, так и за счет углубления. Парциальное давление СО2 в артериальной крови во время мышечной работы не изменяется или незначительно увеличивается. При тяжелой мышечной работе оно меньше, чем при легкой работе. Возбудимость дыхательного центра к действию СО2 во время мышечной работы не увеличивается. Эти факты доказывают, что ведущая роль в увеличении дыхания при мышечной работе принад-лежит не гуморальному, а рефлекторному механизму. Ведущая роль в изменениях дыхания при мышечной работе принадлежит корригирующим влияниям больших полушарий головного мозга и условным рефлексам, главным образом при действии словесных раздражителей. Условные рефлексы на дыхательный аппарат тем больше выражены, чем ближе начало выполнения трудовых или физических упражнений изменения дыхания во время речи обусловлены только нервными влияниями. Рефлекторная регуляция дыхания в соответствии с интенсив-ностью мышечной работы связана с изменениями физико-хими-ческих свойств крови, влияющих на возбудимость дыхательного центра. Максимальный уровень вентиляции легких ограничен пределом возбудимости дыхательного центра. Превышение этого предела при чрезмерном увеличении притока к нему афферентных импульсов уменьшает вентиляцию легких. Легочная вентиляция обычно увеличивается параллельно интенсивности физической работы. Чем напряженнее работа, тем больше воздуха проходит через легкие и тем больше газообмен. При гимнастических упражнениях вентиляция легких увели-чивается в 2-7 раз, а при беге на средние дистанции – даже до 20 раз. При интенсивной длительной мышечной работе, беге, езде на велосипеде, плавании нередко через некоторое время наступает очень сильная одышка, затруднение дыхания и ощущение стесне-ния в груди, учащаются сердцебиения, повышается кровяное давление («мертвая точка»). Это состояние преодолевается волевым усилием, следовательно, импульсами с больших полушарий и тогда исчезает ощущение стеснения в груди и удушье, а самочувствие улучшается. Дыхание становится равным и спокойным, значи-тельно повышается работоспособность (второе дыхание»). При «мертвой точке» в крови накапливаются углекислота и молочная кислота и повышается концентрация водородных ионов. Это вызывает дальнейшее углубление дыхания резко усиленную вентиляцию легких, которая сопровождается увеличением выде-ления СО2 в выдыхаемый воздух и падением концентрации водородных ионов, то же наступает и благодаря увеличенному выделению молочной кислоты при потоотделений в время “второго дыхания”. Усиленная вентиляция легких перед интенсивной мышечной работой наступление «мертвой точки». При подъеме тяжестей, лазании, гребле и других физических упражнениях происходит натуживание – закрытие голосовой щели после вдоха и задержка дыхания при напряжении выдыхательной мускулатуры, что приводит к повышению давления в легких и сужению легочных сосудов. Потребление кислорода в покое и при физической нагрузке В покое человек потребляет 2,5-3 л кислорода в минуту. При мышечной работе значительно увеличивается поглощение кровью кислорода. Максимальная величина поглощения кислорода в минуту («кислородный потолок») у нетренированных не больше 3-3,5 л в минуту, а у хорошо тренированных доходит до 4,5-5,5 л в минуту. Так как при интенсивной мышечной работе потребность в кислороде увеличивается в 10-15 даже 20-25 раз, то образуется «кислородный долг» который погашается после работы в восста-новительном периоде. Кислородный долг – это потребление недос-тающего кислорода после работы. Кислородный долг зависит от тренированности. Он характеризуется способностью выполнять мышечную деятельность при недостаточном снабжений организма кислородом. Кислородный долг возникает потому, что при работе помимо окислительных процессов, сокращаются химические реакции без участия кислорода, анаэробные процессы. При этих анаэробных реакциях возникают химические соединения, которые могут накопиться и временно находиться в каком-то количестве в организме, с тем, чтобы подвергнуться окислению после работы. Усиленное окисление после работы недоокисленных веществ объясняет, почему в течение какого-то времени, иногда значитель-ного, человек после работы усиленно дышит. Сумма кислородного долга и кислорода, потребляемого во время работы, называется кислородным запросом. В зависимости от особенностей мышечной работы и функциональных возможностей организма кислородный запрос не всегда полностью удовлетворяется в процессе выпол-нения работы. Контрольные вопросы В чем состоит сущность и значение дыхания? Каковы фазы дыхательного цикла? Каков механизм транспорта кислорода кровью? Как осуществляется транспорт углекислого газа кровью? Как доказать гуморальную регуляцию активности дыха-тельного центра? Как изменяется функциональная активность системы дыхания при физической нагрузке? Лекция 4 |