Лекции по общей физиологии. 2курс_лекции_по_общей_физиологии. Введение в общую физиологию
 Скачать 6.53 Mb. Скачать 6.53 Mb.
|

|
Тема: “Нервно-мышечная физиология” ПЛАН: Возбудимые ткани и их свойства Нервные волокна, их виды и свойства. Возбуждение, его распространение и передача его через синапс. Понятие о двигательном аппарате и о двигательной еди-нице. Механизм мышечного сокращения и обеспечение его энергией. Типы, формы и режимы мышечного сокращения. Физиологические свойства скелетных и гладких мышц. Электромиограмма и ее показатели. Среди всех тканей организма особо выделяют возбудимые ткани (нервную, мышечную и железистую). Реакция которых на раздражение связана с возникновением специальных форм актив-ности, т.е. процессом возбуждения. Основными функциональными характеристиками возбуди-мых тканей являются раздражимость, возбудимость, возбуждение, лабильность и функциональная подвижность, рефрактерности, торможение, парабиоз. Способность организма, его органов и тканей изменять обмен веществ в ответ на раздражение называется раздражимостью. Возбудимость – свойство возбудимых тканей отвечать на раздражение специфическим процессам возбуждения. Этот процесс включает электрические, ионные, химические и тепловые измене-ния. Внешнее проявление процесса возбуждения: в нервных клет-ках – только импульсы возбуждения; в мышечных – импульсы возбуждения сокращение, а в железистых – выделение определен-ных веществ. Возбуждение ткани представляет собой переход из состояния физиологического покоя в дейтельное состояние. Возбудимые ткани не всегда отвечают на действие раздра-жителя процессом возбуждения, иногда на действие раздражителя наблюдается замедление или прекращение текущей деятельности. Это объясняется процессом торможения. Торможение – это активный процесс который возникает при действии раздражителя. Процессы возбуждения и торможения одних и тех структур находятся в постоянном взаимодействии и определяют функцио-нальную подвижность ткани. Функциональная подвижность или лабильность – это ско-рость перехода из состояния покоя в состояние возбуждения и выхода из него. В одних клетках и тканях возбуждение протекает быстро, а в других медленно. Она может повышаться под влиянием раздражений, тренировки, особенно у спортсменов, при развитии качества быстроты. Во время возбуждения возбудимость ткани изменяется и она не способна реагировать на действие сверх сильных раздражителей. Такое изменение возбудимости ткани обозначается как рефрак-терность, т.е. изменение возбуждения. Изменение возбудимости ткани при их возбуждении проте-кает 2 фазно: 1. Абсолютная фаза рефрактерности; 2. Относи-тельная фаза рефрактерности. Абсолютная фаза рефрактерности, т.е. период полной невозбудимости. Она связана с процессом деполяризации, т.е. с процессом возникновения возбуждения и снижения возбудимости. Относительная фаза связана с процессом утомления, т.е. периодом восстановления возбудимости. Свойства нервов и мышц парабиоз – это состояние нерва на грани жизни и смерти. Процесс развития и углубления парабиоза, по описанию Введенского характеризуется 3 стадиями: 1 – уравнительная. В этой стадии мышца и на слабые и на сильные раздражения нерва дает сокращения одинаковой величины. 2 – парадоксальная, стадия наиболее характерна для пара-биоза. В этой стадии сильные раздражения нерва, вызывают в мышце более низкий тетанус, чем слабые раздражения. А при углублении парабиоза не вызывают никакой ответной реакции, в то время как слабое раздражение дает тетанус. 3 – тормозная, стадия выражается в том, когда и слабые и очень сильные раздражения нерва не вызывают тетанус. В этой стадии наблюдается полная рефрактерность, когда измененный нерв временно потерял способность функционировать, но он еще жив, т.к. при прекращении постоянного действия (химического раздражителя – алкоголь) раздражителя его физиологические свойства восстанавливаются. Таким образом, парабиотическому состоянию нерва и мышц характерно снижение возбудимости и лабильности. Парабиоз есть форма приспособительной реакции живых образований на разнообразные воздействия. Нервные волокна – отростки нейронов длинные – аксоны и короткие – дендриты. Длина нервных волокон более 1 м, толщина волокна одинакова по всей длине и равна нескольким микрометрам. В середине его проходит осевой цилиндр, содержащий нейроплазму и нейрофибриллы. Снаружи нервное волокно покрыто шванновской оболочкой, в которой находится липидное вещество миелин. Нервные волокна, содержащие миелин, называются мякотными, а не содержащие миелин – безмякотными. Большинство нервных волокон мякотные. Наружный слой шванновской оболочки уплотняется и образует тонкую пленку – нейрилемму. В миелиновой оболочке нервных волокон есть прерванные участки – перехваты Ранвье, в которых осевой цилиндр покрыт только нейрилеммой. По строению и функциям периферические мякотные нервные волокна делятся на 3 основные группы: Волокна группы А толстые, мякотные, диаметром от 4 до 20 мкм. К этой группе принадлежат эфферентные волокна скелетной мускулатуры и афферентные волокна, проводящие возбуждение от рецепторов осязания, давления и мышечно-суставной чувствительности. Волокна группы Втонкие, миелиневые волокна диаметром меньше 3 мкм. В эту группу входят афферентные волокна и волокна вегетативной нервной системы. Волокна группы Спокрыты очень тонкой миелиневой оболочкой диаметром меньше 2 мкм. К ним относятся афферентные волокна болевой и температурной чувствительности. В эту группу входят также безмякотные волокна диаметром от 1 до 1,3 мкм. Деятельность возбудимых тканей, а именно нервной ткани является специфичной т.к. деятельным состоянием нерва в основном является их возбуждение. Процесс возбуждения нерва и мышц сопровождается возникновением электрического потенциала. Генерация электрического потенциала возбудимых тканей осуществляется на основе ионных механизмов. Электрические потенциалы проявляются и распространяются в 2-х формах: Локальный (местный) потенциал, который не распро-страняется по мембране нервных волокон. Потенциал действия или распространяющиеся возбуж-дение. Нервные импульсы по волокнам проводятся в виде импульсов прерывисто и непрерывисто. Скорость проведения нервных им-пульсов зависит от диаметра, от сопротивления мембраны и емкости мембраны. Возбуждение в нервных волокнах проводится с большой скоростью в мякотных нервных волокнах, т.к. нервные импульсы проводятся по прерванным участкам т.е. прерывисто по перехватам Ранвье. В немиелинизированных волокнах импульсы проводятся непрерывно. При таком проведении импульса быстро затухают и скорость невелика. Взаимодействие нейронов между собой происходит через специальные образования – синапс. Они образуются концевыми разветвлениями нейрона на теле или отростках другого нейрона. Чем больше синапсов на нервной клетке, тем больше она воспринимает различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия в разно-образных реакциях организма. В структуре синапса различают 3 элемента: Пресинаптическую мембрану, образованную утолщением мембраны конечной веточки аксона; Синаптическая щель между пре и постсинаптической мембранами; Постсинаптическую мембрану – покрывающей нейроны и другие клетки. В большинстве случаев передача возбуждения одного нейрона на другой осуществляется химическим путем. В преси-наптической части имеются синаптические пузырьки, которые содержат специальные вещества – медиаторы или посредники. Ими могут быть ацетилхолин, норадреналин, некоторые аминокислоты и др. Приходящие в окончание аксона нервные импульсы вызывают опорожнение синаптических пузырьков и выведение медиатора в синаптическую щель. По приходу импульса в результате повышения температуры и хаотического движения пузырки лопаются и ацетилхолил вливается в синаптическую щель. Он взаимодействует с холинорецепторами постсинаптической мембраны и приводит к изменению его структуры. В результате этого проницаемость постсинаптической мембраны для ионов Na+ значительно возрастает и она деполя-ризуется. Деполяризация постсинаптической мембраны проявляется в виде электрического ответа постсинаптического потенциала. Как только он достигает критического уровня возникает потенциал действия, который распространяется по всей мышце и она сокращается. В приспособлении организма человека и животных к постоянно меняющимся условиям внешней среды, важное место среди которых занимают дыхательные процессы. Перемещение в пространстве, нахождение пищи, трудовая деятельность невозмож-ны без мышечной работы. Двигательные функции выполняются специальной системой органов и тканей получившей название двигательного аппарата. Двигательный аппарат состоит из сочленных костей скелета и сложной поперечнополосатых мышц, приводящих в движение костные рычаги. К двигательному аппарату также относятся мотонейроны и их аксоны, а также нервно-мышечные синапсы. Скелетные мышцы позвоночных животных снабжаются двигательными нервными волокнами нейронов, находящихся в передних рогах спинного мозга. Каждое нервное волокно, иннер-вирующее группу мышечных волокон, называется двига-тельной единицей (рис.13). 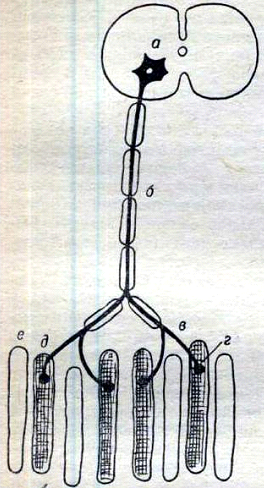 Рис. 13. Схема строения двигательной единицы: а – тело двигательного нейрона; б – двигательное нервное волокно; в – его разветвления; г – нервно-мышечный синапс; д – мышечные волокна, иннервируемые данным нейроном. Различают двигательные единицы, участвующие в фазных движениях (сокращениях и расслаблениях) и в длительных напряжениях мышц. В скелетных мышцах содержаться обе группы волокон. Фазные единицы разделяют на быстрые и медленные, в которых скорость проведения возбуждения в несколько раз меньше, чем в быстрых, а его возникновение и продолжительность сокращения больше. В мышцах человека осуществляются быстрые и точные движения, например в глазных мышцах, в одну двига-тельную единицу входит 3-6, а в мышцах пальцев рук 10-25 мышечных волокон. В мышцах, производящих медленные движе-ния регуляции позы человека, количество мышечных волокон в одной двигательной единице доходит до 2000-3000, в икроножной мышце составляет примерно 2000. Различают малую и большую двигательную единицу. Малая двигательная единица включает маленький мотоней-рон с тонким аксоном, который имеет небольшое число концевых веточек, иннервирующий небольшое число мышечных волокон. Малые двигательные единицы входят в состав всех мелких мышц лицевой мускулатуры пальцев рук, ног, кистей и частично в состав больших мышц туловища и конечностей. Большая двигательная единица включает крупный мотоней-рон с толстым аксоном, который образует большое число концевых веточек в мышце и иннервирует большое число мышечных воло-кон. Большие двигательные единицы входят в состав больших мышц туловища и конечностей. Иннервируемые мотонейронами мышечные волокна содержат миофибриллы, гранулы гликогена капельки жира и др. и сарко-плазматический ретикулум. Миофибрилы представляют собой сократительный аппарат мышечного волокна. В каждом мышечном волокне содержится 1000 более миофибрилл. Диаметр каждой миофибриллы 3 мкм и состоит из толстых и тонких белковых нитей. Толстые белковые нити называются миозинами, тонкие – актиновыми нитями. В тонких нитях содержится 2 вида белка – тропомиозин, тропонин, которые играют существенную роль в сокращении и расслаблении мышц. В естественных условиях организма стимулом к сокращению является потенциал действия. Промежуточным звеном между деполяризацией и началом мышечного сокращения является проникновение ионов калия в миофибриллы. В состоянии покоя ионы калия находятся в цистерне саркоплазматического ретику-лума. Освободившиеся ионы калия из цистерн саркоплазма-тического ретикулума связываются с тропонином и тропомиозином миофибрилл. В результате чего актиновые и миозиновые нити скользят между собой, и проходит сокращение мышц. Такой механизм сокращения мышц объясняется теорией скольжения. Сокращение мышечных волокон происходит с затра-той энергии. Источником энергии для мышечного сокращения является АТФ. При отсутствии АТФ не происходит сокращение мышц т.к. актиновые нити не могут скользить вдоль миозиновых нитей. Энергия требуемая для сокращения мышц выделяется при расщеплении АТФ с участием миозинового фермента. При этом образуется АДФ. При расщеплении 1 молекулы АТФ выделяется около 10 ккал свободной энергии. Расщепление АТФ происходит с определенной скоростью. Скорость расщепления зависит от выполняемой работы мышц. При длительном сокращении мышц для поддержания определенной позы или положения тела необходимо постоянное восстановление АТФ, с такой скоростью какой она расщепляется. При выполнении спортивных упражнений, которые требуют вынос-ливости, скорость расщепления АТФ возрастает в 100 и более раз. Восстановление АТФ происходит 2 путями: анаэробный путь (без участия кислорода); аэробный путь (с участием кислорода). Анаэробный путь – это расщепление гликогена посредством гликолиза. Такой путь восстановления можно наблюдать например при фенальном рывке во время соревнований по бегу. При расщеплении гликогена АТФ восстанавливается в 2-3 раза быстрее и выделяется энергия также в 2-3 раза больше. Аэробный путь восстановления АТФ происходит за счет реакции фосфорилирования с потреблением кислорода. До аэроб-ного образования АТФ. Её восстановление происходит за счет АДФ и креатинфосфата. В зависимости от условий, в которых происходит мышечное сокращение, различают 2 его типа – изотоническое и изомет-рическое. Изотоническим называют такое сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным. Слово «изотонический» обозначает постоянный тонус, т.е. постоянное напряжение. Изометрическим называют такое сокращение, при котором мышца укоротиться не может, меняется лишь ее напряжение: сперва возрастает потом падает. Слово «изометрический» обозна-чает «постоянный по длине». Естественные сокращения мышц в организме никогда не бывают чисто изотоническими или изометрическими, т.к. мышцы, поднимая груз (например, сгибая конечность в суставе) укора-чиваются и вместе с тем, растягивая антагонические мышцы меняют свое напряжение. Такой тип сокращения называют ауксо-тонический. В этом типе протекает работа мышц при выполнении трудовых, спортивных и других двигательных актов. На одиночное раздражение мышца отвечает одиночным сокращением, т.е. кратковременно укорачивается утолщаясь и снова расслабляется. Продолжительность и высота одиночного сокращения зависят прежде всего от функционального состояния мышцы и силы раздражения. В условиях организма в норме одиночное сокращение имеется только у сердечной мышцы. Одиночное сокращение состоит из 3 фаз: 1. латентный период – 2,5 м/сек; 2. фаза сокращения – 50 м/сек; 3. фаза расслабления – 50 м/сек. Факторами, влияющими на функциональное состояние мышцы, являются развитие утомление и температура и т.д. В естественных условиях в организме скелетная мышца получает обычно из нервной системы не одиночные раздражения, а ряд быстро следующих друг за другом нервных импульсов. Под влиянием ритмических раздражений наступает сильное и дли-тельное укорочение мышцы. Такое сокращение называется тетани-ческим сокращением или тетанусом. Тетанические сокращения мышцы представляют собой ре-зультат суммации одиночных сокращений. Для искусственного воспроизведения тетануса на мышцу действуют большим числом раздражений, следующих друг за другом с такой частотой при которой происходит суммация. При относительно малой частоте наступает зубчатый тетанус, при большой частоте – гладкий тетанус. Зубчатый тетанус возникает, если каждое последующее раздражение попадает в фазу расслабления. Гладкий тетанус возникает при совпадении раздражения с фазой укорочения мышц. Различают следующие формы мышечного сокращения: дина-мическое и статическое. Динамическое сокращение наблюдается при последова-тельном чередовании сокращений и расслабления мышц. Статическое сокращение наблюдается когда мышца дли-тельно сокращена без изменения ее длины. Основными физиологическими свойствами (скелетных и гладких) мышц являются: 1) их возбудимость, 2) проводимость, 3) сократимость, 4) лабильность, 5) пластичность, 6) развивать напряжение при сокращении, 7) автоматия Гладкая мышца в отличие от скелетной обладает большой пластичностью т.е. способностью сохранять приданную растя-жением длину без изменения напряжения. Скелетная мышца как резина, после снятия растягивающей её груза, сразу укорачивается. Гладкая мышца после снятия груза остается растянутой до тех пор, пока под влиянием другого какого-либо раздражения не возникает ее активное сокращение. Свойство пластичности имеет большое значение для нормаль-ной деятельности гладких мышц стенок полых органов, например мочевого пузырь: благодаря пластичности гладкой мускулатуры стенок пузыря давление внутри него относительно мало изменяется при разной степени его наполнения. Гладкие мышцы менее возбудимы, чем скелетные: их пороги раздражения выше, а хронаксия длиннее. Потенциалы действия (ПД) большинства гладкомышечных волокон имеют малую амплитуду (порядка 60 мВ ПД вместо 120 мВ ПД в скелетных мышечных волокнах) и большую продолжительность до 1-3 секунд. Скорость проведения возбуждения варьирует в разных волокнах от нескольких миллиметров до нескольких сантиметров в секунду. Характерной особенностью гладких мышц в отличие их от скелетных является способность к спонтанной автоматической деятельности. Спонтанные сокращения можно наблюдать при исследовании гладких мышц желудка, кишок, желчного пузыря, мочеточников и ряда других гладкомышечных органов. Автоматия гладких мышц регулируется нервными элементами, которые находятся в самих мышцах. Спонтанные сокращения гладких мышц происходит в результате медленно развивающейся деполяризации мембраны после каждого потенциала действия. В покое, вне работы, мышцы не являются полностью расслабленными, а сохраняют некоторое напряжение, называемое тонусом. Внешним выражением тонуса является .определенная степень упругости мышц. Например, тоническое напряжение мышц шеи поддерживает положение головы в пространстве, тоническое напряжение мышц туловища и ног – положение тела при стоянии. Электрофизиологические исследования показывают, что тонус скелетных мышц связан с поступлением к мышце отдельно следующих друг за другом с большим интервалом нервных им-пульсов, возбуждающих попеременно различные мышечные волокна. Эти импульсы возникают в мотонейронах спинного мозга, активность которых в свою очередь поддерживается импульсами, исходящими как из вышележащих нервных центров, так и с пери-ферии от рецепторов растяжения (“мышечных веретен”), находя-щиxся в самих мышцах. О рефлекторной природе тонуса скелетных мышц свиде-тельствует тот факт, что перерезка задних корешков спинного мозга, по которым чувствительные импульсы от мышечных веретен поступают в спинной мозг, приводит к полному расслаблению мышцы. У человека тонус мышц может регулироваться произвольно – по желанию человека, может почти полностью расслабить мышцы или же несколько напрячь их не совершая при этом движения. Для измерения силы мышцы определяют тот максимальный груз, который она в состоянии поднять. Эта сила может быть очень велика. Установлено, что собака мышцами челюсти может поднять груз превышающий вес ее тела в 8,3 раза. О силе икроножных мышц человека судят по величине груза, положенного ему на плечи, с которым он в состоянии приподняться на носки. Сила мышцы не зависит от ее длины, а зависит от ее толщи-ны, от физиологического поперечника т.е. от количества мышечных волокон, приходящихся на наибольшую площадь ее поперечного сечения. Физиологическим поперечником называется площадь сечения всех мышечных волокон. У перистых и полуперистых мышц этот поперечник больше анатомического. У веретенообразных и параллельных мышц физиологический поперечник совпадает с анатомическим. Поэтому наиболее сильные перистые мышцы, затем полуперистые, веретенообразные и наиболее слабые мышцы с параллельным ходом волокон (рис. 14). 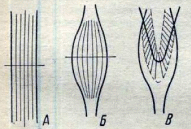 Рис. 14. Физиологический и анатомический поперечники в параллельно волокнистой (А), веретенообразной (Б) и перистой (В) мышцах Сила мышц зависит от 1) сократительной способности мышечных волокон (суммы сил мышечных волокон); 2) от количества мышечных волокон в мышце и функциональных единиц; 3) от условий взаимодействия мышцы с костями скелета. Сила мышц повышается при тренировке, снижается при голодании и утомлении. Вначале она увеличивается с возрастом, а затем к старости уменьшается. Различают абсолютную и относительную силу мышц. Вбсолютная мышечная сила – это отношение максимальной силы мышцы к её физиологическому поперечнику. Абсолютная сила, выраженная в кг на 1 см2 икроножной мышцы человека равна 5,9; сгибателя плеча – 8,1; жевательной мышцы – 10. Относительная сила мышцы высчитывается следующим образом. Определив абсолютную силу в кг или Ньютонах, делят ее на число квадратных см поперечного сечения мышцы. Это позволяет сравнить силу разных мышц одного и того же организма. Силу одноименных мышц разных организмов, а также изменения силы одной и той же мышцы данного организма в зависимости от сдвигов ее функционального состояния. Относительная сила скелетной мышцы лягушки 2-3 кг, разгибателя шеи человека – 9 кг, жевательной мышцы – 10 кг, двуглавой мышцы плеча – 11 кг, трехглавой мышцы плеча – 17 кг. Анализ работы мышц при совершении двигательного акта осуществляется с помощью метода электромиографии т.е. регис-трирует потенциалы действия, возникающих в мышце при возбуждении. При электромиографическом изучении движений обычно используют накожные электроды, укрепляемые над исследуемой мышцей. Многоканальный электромиограф дает возможность одновременно записывать электромиограммы нескольких мышц. Между амплитудой зубцов электромиограммы и развиваемой мышцей силой существует примерно линейная зависимость. Поэтому электромиограммы, в сопоставлении с регистрируемыми синхронно механограммами, позволяют судить о силе сокращения мышц и о распределении их активности в последовательных фазах двигательного акта. Метод электромиографии широко применяется в физиологии спорта и медицине для оценки состояния двигательного аппарата и диагностики ряда заболеваний. Электроды различных типов позволяют отводить внеклеточно потенциалы целой мышцы, отдельных двигательных единиц и даже отдельных волокон. Электромиография позволяет выявлять разнообразные нарушения иннервации мышц и их управления ЦНС. Контрольные вопросы Назовите физиологические свойства нервной и мышечной ткани? Дайте краткую характеристику возбудимости, проводи-мости, рефрактерности, лабильности? Какова причина возникновения потенциала покоя нерва? Какие существуют виды нервных волокон в зависимости от особенностей их строения? Как возникает потенциал действия? Что такое синапс? Что такое парабиоз и каковы стадии его развития? Перечислите физиологические свойства скелетных мышц и дайте их характеристику? Перечислите физиологические особенности гладких мышц и дайте их характеристику? ЛЕКЦИЯ 10 |