Зачем врачу нужна биологическая химия
 Скачать 6.47 Mb. Скачать 6.47 Mb.
|

Газообмен происходит по градиенту концентрацийОбмен кислорода и углекислого газа в тканяхВ тканях диффундирующий в кровь из клеток СО2 большей частью (около 90%) по градиенту концентрации попадает в эритроциты. Движущей силой этого процесса является быстрая, постоянно идущая реакция превращения его в угольную кислоту при участии фермента карбоангидразы. Угольная кислота диссоциирует и подкисляет содержимое эритроцита, что улучшает отдачу оксигемоглобином кислорода (Эффект Бора). 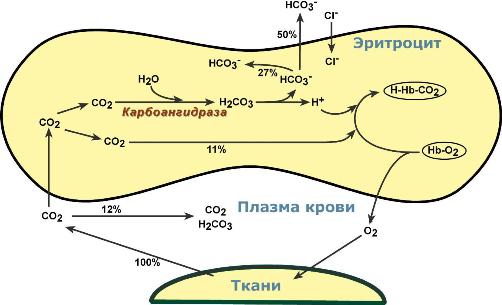 Реакции газообмена в капиллярах тканейОдновременно с концевыми NH2-группами β-цепей гемоглобина связывается 10-12% карбонат-иона с образованием карбаминогемоглобина(H-HbCO2). Остальные бикарбонаты выходят в плазму крови в обмен на ионы хлора (гипохлоремический сдвиг). Обмен кислорода и углекислого газа в легкихВ легочных капиллярах имеется относительно низкая концентрация углекислого газа в альвеолярном воздухе: 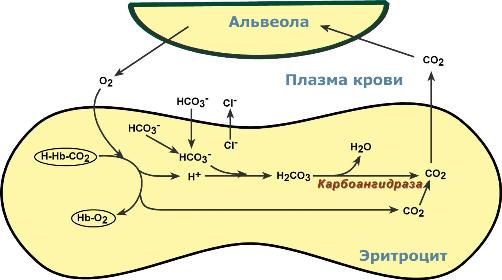 Реакции газообмена в легочных капиллярах
Гем требуется для многих ферментовГем является небелковой частью многих гемопротеинов:
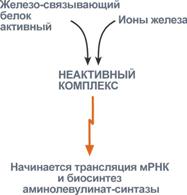
Строение и синтез гемаСинтез гема в основном идет в предшественниках эритроцитов, клетках печени, почек, слизистой кишечника, и в остальных тканях. Первая реакция синтеза с участием δ-аминолевулинат-синтазы (греч. δ - "дельта") происходит в митохондриях. Следующая реакция при участии аминолевулинатдегидратазы(порфобилиноген-синтазы) протекает в цитозоле. 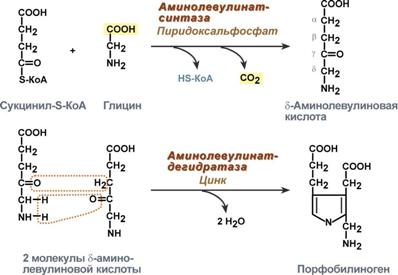 Синтез порфобилиногенаПосле синтеза порфобилиногена четыре его молекулы конденсируются в тетрапиррол. Различают два вида тетрапирролов - уропорфириноген типа I и уропорфириноген типа III. В синтезе обоих видов порфиринов принимает участие уропорфириноген I-синтаза, в образования уропорфириногена III дополнительно принимает участие фермент уропорфириноген III-косинтаза. 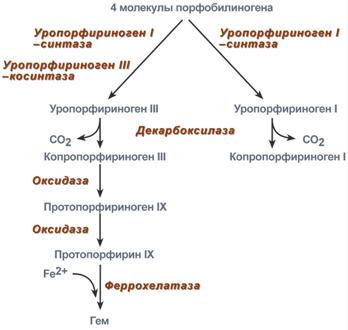 Синтез гема из порфобилиногенаДалее уропорфириногены превращаются в соответствующие копропорфириногены. Копропорфириноген III окисляется в протопорфириноген IX и далее в протопорфирин IX. Последний после связывания с железом образует гем, реакцию катализирует феррохелатаза(гемсинтаза). Регуляция синтеза гема1. Скорость синтеза глобиновых цепей зависит от наличия гема, он ускоряет биосинтез "своих" белков. 2. Основным регуляторным ферментом синтеза гема является аминолевулинатсинтаза.
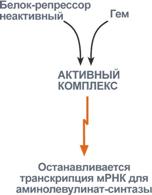
3. Положительным модулятором аминолевулинатсинтазы служит гипоксиятканей, которая в эритропоэтических тканях индуцирует синтез фермента. 4. В печени повышение активности аминолевулинатсинтазы вызывают соединения, усиливающие работу микросомальной системы окисления (жирорастворимые токсины, стероиды) – при этом возрастает потребление гема для образования цитохрома Р450, что снижает внутриклеточную концентрацию свободного гема. В результате происходит дерепрессия синтеза фермента. |