Зачем врачу нужна биологическая химия
 Скачать 6.47 Mb. Скачать 6.47 Mb.
|

Жиры синтезируются только при наличии энергииРеакции биосинтеза липидов могут идти в гладкой эндоплазматической сети клеток всех органов. Субстратом для синтеза жиров de novo является глюкоза. Как известно, попадая в клетку, глюкоза превращается в гликоген, пентозы и окисляется до пировиноградной кислоты. При высоком поступлении глюкоза используется для синтеза гликогена, но этот вариант ограничивается объемом клетки. Поэтому глюкоза "проваливается" в гликолиз и превращается в пируватлибо напрямую, либо через пентозофосфатный шунт. Во втором случае образуется НАДФН, который понадобится впоследствии для синтеза жирных кислот. 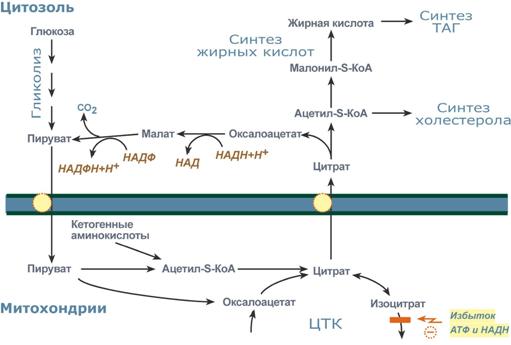 Общая схема биосинтеза триацилглицеролов и холестерола из глюкозыПируват переходит в митохондрии и декарбоксилируется в ацетил-SКоА и вступает в ЦТК. Однако в состояниипокоя, при отдыхе, при наличии избыточного количества энергиив клетке реакции ЦТК (в частности, изоцитратдегидрогеназная реакция) блокируютсяизбытком АТФ и НАДН. В результате накапливается первый метаболит ЦТК – цитрат. По градиенту концентрации он перемещается в цитозоль, расщепляется с образованием ацетил-SКоА, который далее используется в биосинтезехолестерола, жирных кислоти триацилглицеролов. Оксалоацетат, также образуемый из цитрата, восстанавливается до яблочной кислоты и возвращается в митохондрии
Синтез жирных кислот не похож на путь их окисленияБиосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покояили после еды. Условно можно выделить 4 этапа биосинтеза: 1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот. 2. Перенос ацетил-SКоА из митохондрий в цитозоль:
Поступающий из митохондрий цитрат в цитозоле расщепляетсяАТФ-цитрат-лиазойдо оксалоацетата и ацетил-SКоА.Оксалоацетат в дальнейшем восстанавливается до малата, и последний либо переходит в митохондрии (малат-аспартатный челнок), либо декарбоксилируется в пируват малик-ферментом ("яблочный" фермент). 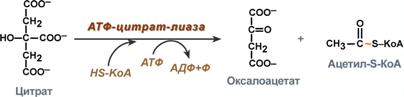 Образование ацетил-SКоА из лимонной кислоты3. Образование малонил-SКоА из ацетил-SКоА. Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов. 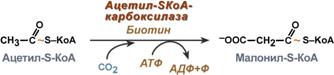 Образование малонил-SКоА из ацетил-SКоА 4. Синтез пальмитиновой кислоты. Осуществляется мультиферментнымкомплексом "синтаза жирных кислот" (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ). Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин (ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН. 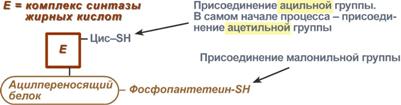 Активные группы синтазы жирных кислотВ первых двух реакциях последовательно присоединяются малонил-SКоА к фосфопантетеину ацил-переносящего белка и ацетил-SКоА к цистеину 3-кетоацилсинтазы. 3-Кетоацилсинтаза катализирует третью реакцию – перенос ацетильной группы на С2 малонила с отщеплением карбоксильной группы. Далее кетогруппа в реакциях восстановления (3-кетоацил-редуктаза), дегидратации (дегидратаза) и опять восстановления (еноил-редуктаза) превращается в метиленовую с образованием насыщенного ацила,связанного с фосфопантетеином. Ацилтрансферазапереносит полученный ацил на цистеин 3-кетоацил-синтазы, к фосфопантетеину присоединяется малонил-SКоА и цикл повторяется 7 раз до образования остатка пальмитиновой кислоты. После этого пальмитиновая кислота отщепляется шестым ферментом комплекса тиоэстеразой. 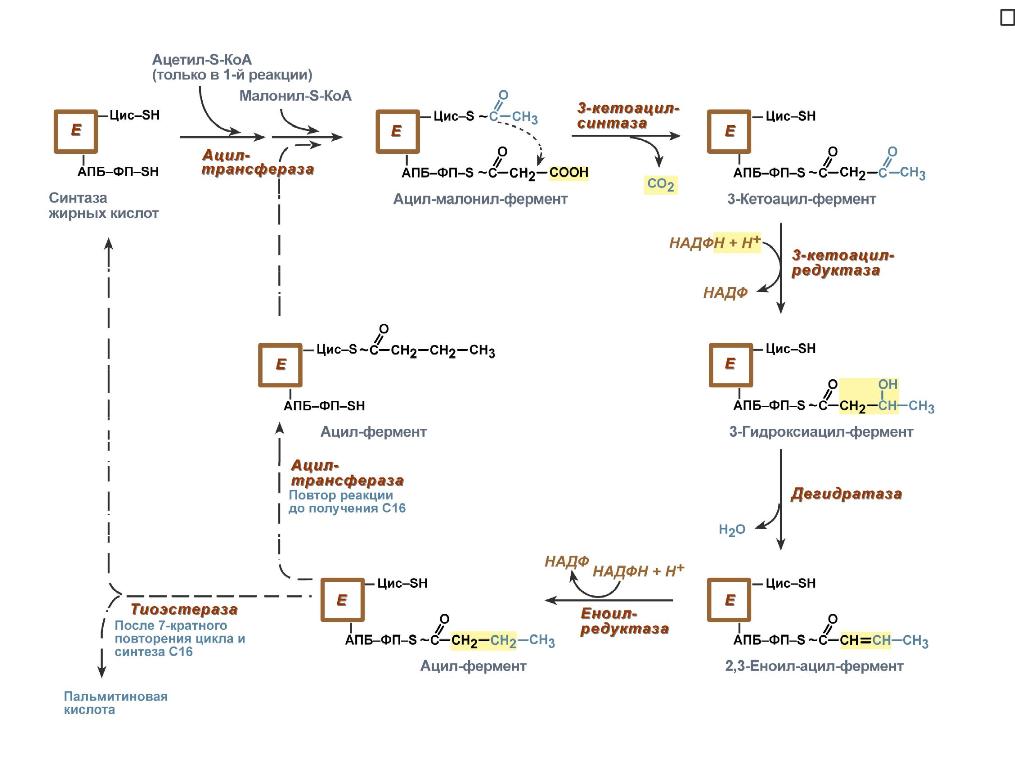 Реакции синтеза жирных кислотУдлинение цепи жирных кислотСинтезированная пальмитиновая кислота при необходимости поступает в эндоплазматический ретикулум или в митохондрии. Здесь с участием малонил-S-КоА и НАДФН цепь удлиняется до С18 или С20. Удлиняться могут и ненасыщенные жирные кислоты (олеиновая, линолевая, линоленовая) с образованием производных эйкозановой кислоты (С20). Но двойная связь животными клетками вводится не далее 9 атома углерода, поэтому ω3- и ω6-полиненасыщенные жирные кислоты синтезируются только из соответствующих предшественников. Например, арахидоновая кислота может образоваться в клетке только при наличии линоленовой или линолевой кислот. При этом линолевая кислота (18:2) дегидрируется до γ-линоленовой (18:3) и удлиняется до эйкозотриеновой кислоты (20:3), последняя далее вновь дегидрируется до арахидоновой кислоты (20:4). Так формируются жирные кислоты ω6 ряда Для образования жирных кислот ω3-ряда, например, тимнодоновой (20:5), необходимо наличие α-линоленовой кислоты (18:3), которая дегидрируется (18:4), удлиняется (20:4) и опять дегидрируется (20:5). |