Зачем врачу нужна биологическая химия
 Скачать 6.47 Mb. Скачать 6.47 Mb.
|

Активность ТАГ-липазы зависит от гормоновПервое, что происходит при использовании нейтрального жира во время голодания и физической нагрузки – это активация фермента, отвечающего за отщепление первой жирной кислоты от триацилглицерола. Фермент называется гормон-чувствительная триацилглицерол-липаза или ТАГ-липаза. Кроме ТАГ-липазы, в адипоцитах имеются еще диацилглицерол-липаза ( ДАГ-липаза) и моноацилглицерол-липаза (МАГ-липаза), активность которых высока и постоянна, однако в покое эта активность не проявляется из-за отсутствия субстрата. Как только в клетке после работы ТАГ-липазы появляются диацилглицеролы начинает работать постоянно активная ДАГ-липаза, продукт ее реакции моноацилглицерол (МАГ) является субстратом для МАГ-липазы. Образующиеся жирные кислоты и глицерол покидают клетку. 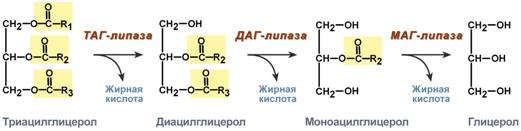 Гидролиз триацилглицеролов липазами жировой клеткиДля регуляции активности ТАГ-липазы обязательно наличие гормонального влияния (адреналин, глюкагон, соматотропин, инсулин и ряд других гормонов). Активация триацилглицерол-липазыГормонзависимая активация ТАГ-липазы адипоцитов адреналиноми глюкагономпроисходит при напряжении организма (голодание, длительная мышечная работа, охлаждение). Активность ТАГ-липазы зависит главным образом от соотношения инсулин / глюкагон. В целом последовательность событий активации липолиза выглядит следующим образом:
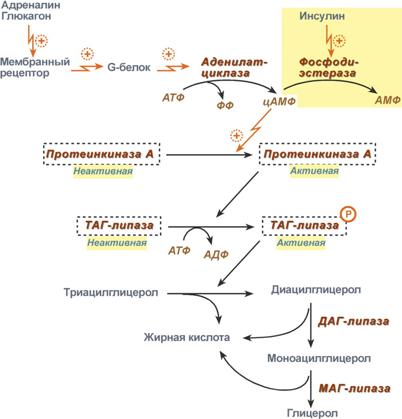 Каскадный механизм активации ТАГ-липазыКроме гормонов, влияющих на активность аденилатциклазы через G-белки, существуют иные механизмы активации. Например, соматотропный гормон увеличивает количество аденилатциклазы, глюкокортикоидыспособствуют синтезу ТАГ-липазы. Снижение активности ТАГ-липазыИнсулинпрепятствует активации липолиза другими гормонами, т.к.
Для сжигания жирных кислот существует свой путьОкисление жирных кислот (β-окисление)Для преобразования энергии, заключенной в жирных кислотах, в энергию связей АТФ существует метаболический путь окисления жирных кислот до СО2 и воды, тесно связанный с циклом трикарбоновых кислот и дыхательной цепью. Этот путь называется β-окисление, т.к. происходит окисление 3-го углеродного атома жирной кислоты (β-положение) в карбоксильную группу, одновременно от кислоты отщепляется ацетильная группа, включающая С1 и С2 исходной жирной кислоты. 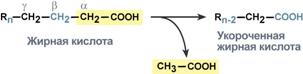 Элементарная схемаβ-окисленияРеакции β-окисления происходят в митохондрияхбольшинства клеток организма (кроме нервных клеток). Для окисления используются жирные кислоты, поступающие в цитозоль из кровиили появляющиеся прилиполизесобственных внутриклеточных ТАГ. Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом: Пальмитоил-SКоА + 7ФАД + 7НАД+ + 7Н2O + 7HS-KoA → 8Ацетил-SКоА + 7ФАДН2 + 7НАДН Этапы окисления жирных кислот1. Прежде, чем проникнуть в матрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-S-КоА. Ацил-S-КоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты. 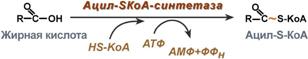 Реакция активации жирной кислоты2. Ацил-S-КоА не способен проходить через митохондриальную мембрану, поэтому существует способ его переноса в комплексе с витаминоподобным веществом карнитином. На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I. 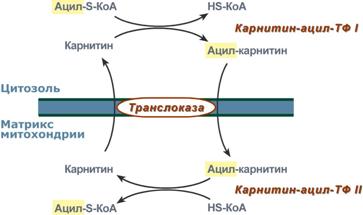 Карнитин-зависимый транспорт жирных кислот в митохондриюКарнитин синтезируется в печени и почках и затем транспортируется в остальные органы. Вовнутриутробномпериоде и в первые годы жизни значение карнитина для организма чрезвычайно важно. Энергообеспечение нервной системы детскогоорганизма и, в частности, головного мозга осуществляется за счет двух параллельных процессов: карнитин-зависимого окисления жирных кислот и аэробного окисления глюкозы. Карнитин необходим для роста головного и спинного мозга, для взаимодействия всех отделов нервной системы, ответственных за движение и взаимодействие мышц. Существуют исследования, связывающие с недостатком карнитина детский церебральный паралич и феномен "смерти в колыбели". 3. После связывания с карнитином жирная кислота переносится через мембрану транслоказой. Здесь на внутренней стороне мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-S-КоА который вступает на путь β-окисления. 4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА. 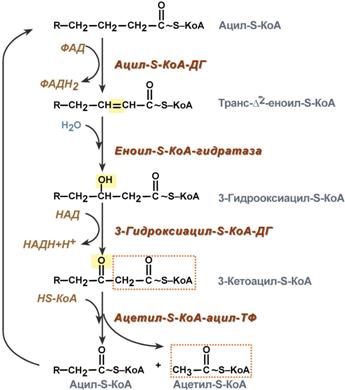 Последовательность реакций β-окисления жирных кислотРасчет энергетического баланса β-окисленияПри расчете количества АТФ, образуемого при β-окислении жирных кислот необходимо учитывать:
Пример. Окисление пальмитиновой кислоты
Таким образом, суммируя, получаем 96+35-2 =129 молекул АТФ образуется при окислении пальмитиновой кислоты. |