Значение биохимии в подготовке врача. Биологическая химия
 Скачать 8.33 Mb. Скачать 8.33 Mb.
|

|
43. Биологические мембраны, строение, функции и общие свойства: жидкостность, поперечная ассиметрия, избирательная проницаемость. Строение и состав мембран Биологические мембраны представляют собой "ансамбли" липидных и белковых молекул, удерживаемых вместе с помощью нековалент-ных взаимодействий. Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолштиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы "растворены" в липидном бислое. 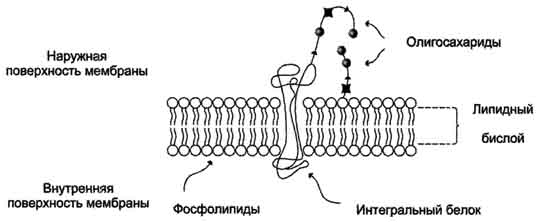 Рис. 5-1. Поперечный разрез плазматической мембраны. 1. Структура и свойства липидов мембран Мембранные липиды - амфифильные (амфипатические) молекулы, т.е. в молекуле есть как гидрофильные группы (полярные "головки"), так и алифатические радикалы (гидрофобные "хвосты"), самопроизвольно формирующие бислой. В большинстве эукариотических клеток они составляют около 30-70% массы мембраны (рис. 5-2). В мембранах присутствуют липиды трёх главных типов - фосфолипиды, гликолипиды и холестерол (холестерин). Липидный состав мембран различен, содержание того или другого липида, по-видимому, определяется разнообразием функций, выполняемых этими липидами в мембранах. Фосфолипиды. Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран - фосфатидилхолины и фосфатидилэтаноламины (рис. 5-3). В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных "головок" (табл. 5-1), так и ацильных остатков (табл. 5-2). Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирно-кислотных остатков. В состав фосфатидил-1инозитол-4,5-бисфосфатов входят в основном ацильные остатки стеариновой или пальмитиновой (по первому положению глицерола) и арахидоновой (по второму положению) жирных кислот. Специфические фосфолипиды внутренней мембраны митохондрий - кардиолипины (дифосфатидилглицеролы), построенные на основе глицерола и двух остатков фосфатид-ной кислоты. Они синтезируются ферментами внутренней мембраны митохондрий и составляют около 22% от всех фосфолипидов мембраны. В плазматических мембранах клеток в значительных количествах содержатся сфингомие-лины (рис. 5-4). Сфингомиелины построены на основе церамида - ацилированного аминоспирта сфингозина. Полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Сфингомиелины - главные липиды миелиновой оболочки нервных волокон. Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа - углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида (рис. 5-5). В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно- или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA). Полярные "головки" гликосфинголипидов находятся на наружной поверхности плазматических мембран. В значительных количествах гликолипиды содержатся в мембранах клеток мозга, эритроцитов, эпителиальных клеток. Ганглиозиды эритроцитов разных индивидуумов различаются строением олигосахаридных цепей, проявляющих антигенные свойства. Холестерол. Холестерол присутствует во всех мембранах животных клеток. Его молекула состоит из жёсткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является "полярной головкой". Для животной клетки среднее молярное отношение холестерол/фосфолипиды равно 0,3-0,4, но в плазматической мембране это соотношение гораздо выше (0,8-0,9). Наличие холестерола в мембранах уменьшает подвижность жирных кислот, снижает латеральную диффузию липи-дов и белков, и поэтому может влиять на функции: мембранных белков. В составе мембран растений холестерола нет, а присутствуют растительные стероиды - ситостерол и стигмастерол. 2. Трансмембранная асимметрия липидов Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам - эту особенность мембран называют трансмембранной (поперечной) асимметрией. Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными "головками" стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную "головку", больше. Фосфатадилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем. Липиды в некоторых биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, т.е. совершают "флип-флоп" (от англ, flip-flop) перескоки (рис. 5-7). Перемещение липидных молекул затрудняют полярные "головки", поэтому липиды, находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость трансмембранной миграции по сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или вообще не совершающими "флип-флоп" перескоки. 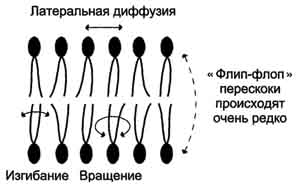 Рис. 5-7. Типы движений липидных молекул в бислое мембран. 3. Жидкостностъ мембран Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот. Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые "изломы" (см. раздел 8). Эти "изломы" препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно и более "текучей". На текучесть мембран также влияют размеры углеводородных "хвостов" липидов, с увеличением длины которых мембрана становится более "текучей". 4. Функции мембранных липидов Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций. Липиды формируют среду для функционирования Мембранных белков, принимающих в ней нативную конформацию. Выделенные из мембран ферменты, лишённые липидного окружения, как правило, не проявляют каталитической активности. Некоторые мембранные липиды - предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфати-дилинозитол-4,5-бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАТ), активатора протеинкиназы С и инозитол-1,4,5-трифосфата (ИФ3) - регулятора кальциевого обмена в клетке (рис. 5-8). ДАГ, ИФ3, протеинкиназа С и Са2+ - участники инозитолфосфатной системы передачи сигнала. Кроме того, некоторые липиды выполняют "якорную" функцию, например к фосфатидил-инозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности клетки. Фосфатидилинозитол с присоединённым к нему олигосахаридом (гликаном) называют фосфатидилинозитолгликаном. Связь белков с этой молекулой (гликаном) осуществляется через фосфоэтаноламин. 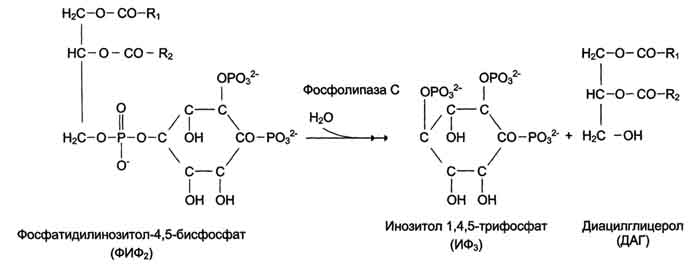 Рис. 5-8. Гидролиз фосфатидилинозитол-4,5-бисфосфата. Пример такого "заякоренного" белка - ацетилхолинэстераза, катализирующая гидролиз ацетилхолина в синаптической щели. Этот фермент фиксируется на поСтеинаптической мембране, ковалентно присоединяясь к фосфатидилино-зитолгликану. Под действием фосфолипазы С может происходить отделение белков от внешней поверхности клетки. Липиды могут быть аллостерическими активаторами мембранных ферментов. Например, р-гидроксибутиратдегидрогеназа, участвующая в окислении кетоновых тел (см. раздел 8), локализована на внутренней мембране митохондрий. Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина. Фермент протеинкиназа С катализирует реакции фосфорилирования белков по аминокислотным остаткам серина и треонина. В неактивной форме протеинкиназа С находится в цитозоле. Однако после стимуляции клетки (повышение в клетке концентрации кальция) фермент быстро активируется ионами кальция и оказывается связанным с мембраной. Функционально активная протеинкиназа С - комплекс, содержащий мономер фермента, молекулу диацилглицерола, один или более ионов Са2+ и четыре молекулы фосфатидилсерина. Креатинкиназа, фермент катализирующий образование макроэргического соединения креа-тинфосфата (см. раздел 9). Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны митохондрий. 44. Липидный состав мембран - фосфолипиды, гликолипиды, холестерин. Белки мембран - интегральные, поверхностные, «заякоренные». Роль отдельных компонентов мембран в формировании структуры и выполнении функций. Липидам принадлежит главная роль в образовании мембран как клеточных структур: пластинчатая, «мембранная» форма и основные физико-химические свойства мембран определяются именно липидами. Основная часть липидов (до 90 %) в мембранах представлена фосфолипидами, гликолипидами и холестерином. Фосфолипиды. В мембранах имеются фосфолипиды двух типов — глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды. Эти липиды являются производными фосфатидной кислоты (диацилглицеринфосфата). В состав фосфолипидов входят жирные кислоты, как насыщенные, так и ненасыщенные. Наиболее распространенные глицерофосфолипиды — это фосфатидилхолины. Их отличительной чертой является наличие в молекуле остатка холина, связанного с фосфорной кислотой. Фосфатидилхолинами называют группу соединений, отличающихся друг от друга природой жирно-кислотных остатков (радикалов R). По такому же типу построены и другие глицерофосфолипиды — фосфатидилэтаноламины и фосфатидилсерины, содержащие соответственно этаноламин и серии. 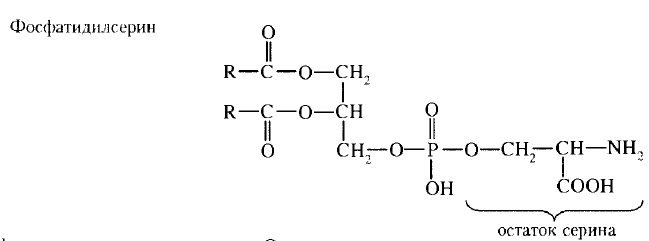 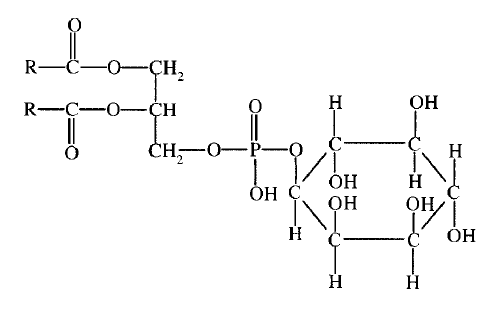 Специальные функции в мембранах выполняет фосфатидилинозитол, в состав которого входит шестиатомный циклический спирт инозитол. Этот фосфолипид участвует в одном из механизмов передачи внешних регуляторных сигналов через клеточную мембрану в клетку. Сфингофосфолипиды (сфингомиелины). В эту группу входят липиды, содержащие аминоспирт сфингозин. Сфингофосфолипиды являются производными церамидов (N-ацилсфингозинов); в остальном сфингофосфолипиды построены сходно с глицерофосфолипидами. 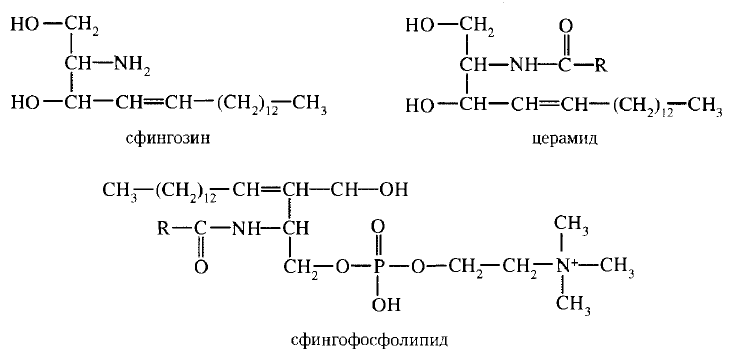 Церамиды — это группа соединений, различающихся по остатку жирной кислоты. В сфингофосфолипидах водород гидроксильной группы у первого углеродного атома церамида замещен на фосфохолин, фосфоэтаноламин или фосфосерин. Гликолипиды представляют собой углеводсодержащие соединения, в которых углеводная часть ковалентно связана с липидной. В мембранах содержатся главным образом углеводные производные церамида (N-ацилсфингозина). Общее название таких гликолипидов — гликозилцерамиды, гликосфинголипиды (их называют также цереброзидами). Напомним, что приставки глюко- или глюкозил- означают глюкозный остаток, а глико- и гликозил- — любой углеводный остаток. Углеводная часть в гликозилцерамидах может быть представлена моносахаридами или олигосахаридами, например остатком лактозы (дисахарид) в лактозилцерамиде. В меньших количествах в гликолипидах обнаруживаются углеводы более сложного строения, образующие разветвленные цепи. Они содержат до 20-30 моносахаридов. Концевые остатки углеводных цепей часто представлены N-ацетилней-раминовой кислотой (девятиуглеродный моносахарид); гликолипиды, содержащие N-ацетилнейраминовую кислоту, называют ганглиозидами. Холестерин — это представитель группы липидов, называемых стероидами. Характерные черты структуры холестерина — наличие тетрациклической группировки и углеводородной разветвленной цепи с восемью углеродными атомами; в третьем положении полициклической части имеется спиртовая группа. 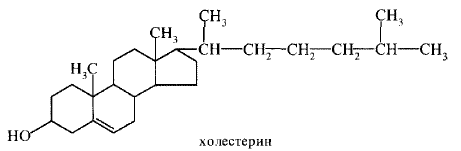 Белки мембран. Мембранные белки, контактирующие с гидрофобной частью липидного бислоя, должны быть амфифильными. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных "головок", обогащены гидрофильными аминокислотными остатками. Белки мембран различаются по своему положению в мембране. Они могут глубоко проникать в липидный бислой или даже пронизывать его - интегральные белки,либо разными способами прикрепляться к мембране - поверхностные белки. Поверхностные белки. Прикрепляются к мембране, взаимодействуя с интегральными белками или поверхностными участками липидного слоя. Белки, образующие комплексы с интегральными белками мембраны. Ряд пищеварительных ферментов, участвующих в гидролизе крахмала и белков, прикрепляется к интегральным белкам мембран микроворсинок кишечника. Примерами таких комплексов могут быть сахаразаизомальтаза и мальтазагликоамилаза. Возможно, связь этих пищеварительных ферментов с мембраной позволяет с высокой скоростью гидролизовать субстраты и усваивать продукты гидролиза клеткой. Белки, связанные с полярными «головками» липидов мембран. Полярные или заряженные домены белковой молекулы могут взаимодействовать с полярными "головками" липидов, образуя ионные и водородные связи. Кроме того, множество растворимых в цитозоле белков при определённых условиях могут связываться с поверхностью мембраны на непродолжительное время. Иногда связывание белка -- необходимое условие проявления ферментативной активности. К таким белкам, например, относят протеинкиназу С, факторы свёртывания крови. Закрепление с помощью мембранного "якоря" "Якорем" может быть неполярный домен белка, построенный из аминокислот с гидрофобными радикалами. Примером такого белка может служить цитохром b5 мембраны ЭР. Этот белок участвует в окислительно-восстановительных реакциях, как переносчик электронов. Роль мембранного "якоря" может выполнять также ковалентно связанный с белком остаток жирной кислоты (миристиновой - С14 или пальмитиновой - С16). Белки, связанные с жирными кислотами, локализованы в основном на внутренней поверхности плазматической мембраны. Миристиновая кислота присоединяется к N-концевому глицину с образованием амидной связи. Пальмитиновая кислота образует тиоэфирную связь с цистеином или сложноэфирную с остатками серина и треонина. Небольшая группа белков может взаимодействовать с наружной поверхностью клетки с помощью ковалентно присоединённого к С-концу белка фосфатидилинозитолгликана. Этот "якорь" - часто единственное связующее звено между белком и мембраной, поэтому при действии фосфолипазы С этот белок отделяется от мембраны. Трансмембранные (интегральные) белки Некоторые из трансмембранных белков пронизывают мембрану один раз (гликофорин), другие имеют несколько участков (доменов), последовательно пересекающих бислой. Трансмембранные домены, пронизывающие бислой, имеют конформацию α-спирали. Полярные остатки аминокислот обращены внутрь глобулы, а неполярные контактируют с мембранными липидами. Такие белки называют "вывернутыми" по сравнению с растворимыми в воде белками, в которых большинство гидрофобных остатков аминокислот спрятано внутрь, а гидрофильные располагаются на поверхности. Радикалы заряженных аминокислот в составе этих доменов лишены заряда и протонированы (-СООН) или депротонированы (-NH2). 45. Механизм переноса веществ через мембраны: простая диффузия, пассивный симпорт и антипорт, активный транспорт, регулируемые каналы. Мембранные рецепторы. Любая молекула может пройти через липидный бислой, однако скорость пассивной диффузиивеществ, т.е. перехода вещества из области с большей концентрацией в область с меньшей, может сильно отличаться. Для некоторых молекул это занимает столь длительное время, что можно говорить об их практической непроницаемости для липидного бислоя мембраны. Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах. Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Малые полярные незаряженные молекулы - СО2, NH3, Н2О, этанол, мочевина - также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана непроницаема. Транспорт таких молекул возможен благодаря наличию в мембранах либо белков, формирующих в липидном слое каналы (поры), заполненные водой, через которые могут проходить вещества определённого размера простой диффузией, либо специфических белков-переносчиков, которые избирательно взаимодействуя с определёнными лигандами, облегчают их перенос через мембрану(облегчённая диффузия). Кроме пассивного транспорта веществ, в клетках есть белки, активно перекачивающие определённые растворённые в воде вещества против их градиента, т.е. из меньшей концентрации в область большей. Этот процесс, называемый активным транспортом, осуществляется всегда с помощью белков-переносчиков и происходит с затратой энергии. |