Значение биохимии в подготовке врача. Биологическая химия
 Скачать 8.33 Mb. Скачать 8.33 Mb.
|

|
Aдгезивные белки К первой группе белков с выраженными адгезивными свойствами относят фибронектин, ламинин, нидоген, фибриллярные коллагены и коллаген IV типа; их относят к белкам "зрелой" соединительной ткани. Фибронектин Фибронектин - один из ключевых белков межклеточного матрикса, неколлагеновый структурный гликопротеин, синтезируемый и выделяемый в межклеточное пространство многими клетками. Он построен из двух идентичных полипептидных цепей, соединённых дисульфидными мостиками у своих С-концов (рис. 15-19). Полипептидная цепь фибронектина содержит 7-8 доменов, на каждом из которых расположены специфические центры для связывания разных веществ. Фибронектин может связывать 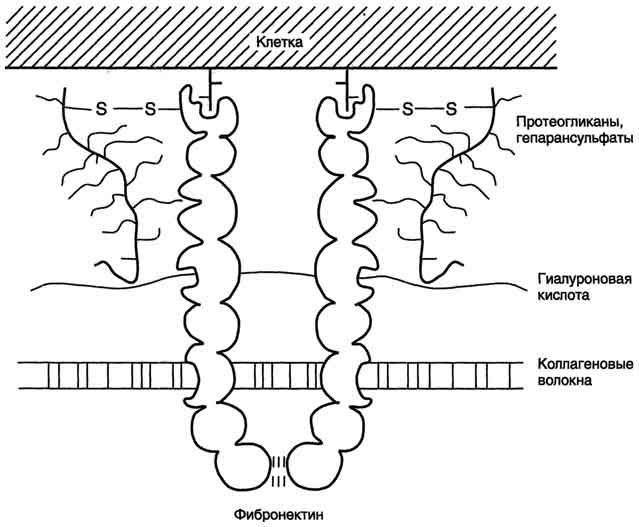 Рис. 15-19. Строение фибронектина. коллаген, протеогликаны, гиалуроновую кислоту, углеводы плазматических мембран, гепарин, фермент трансглутаминазу. Благодаря своей структуре фибронектин может выполнять интегрирующую роль в организации межклеточного вещества, а также способствовать адгезии клеток. Существует несколько форм фибронектина, которые синтезируются разными клетками. Растворимый, или плазменный, фибронектин синтезируется гепатоцитами. Нерастворимый, или тканевый фибронектин синтезируется в основном фибробластами или эндотелиоцитами, глиоцитами и эпителиальными клетками. Обе формы фибронектина вовлекаются в разнообразные процессы: способствуют адгезии и распространению эпителиальных и мезенхимальных клеток, стимулируют пролиферацию и миграцию эмбриональных и опухолевых клеток, контролируют дифференцировку и поддержание цитоскелета клеток, активно участвуют в воспалительных и репаративных процессах. Это связано с тем, что каждая субъединица фибронектина содержит последовательность Арг-Гли-Асп (RGD), с помощью которой он может присоединяться к клеточным рецепторам (интегринам). Эти рецепторы опосредованно взаимодействуют с актиновыми микрофиламентами, которые находятся в цитозоле. В этом процессе участвуют так называемые белки прикрепления (от англ. attach - прикреплять proteins): талин, винкулин, α-актинин (рис. 15-20). С помощью таких белок-белковых взаимодействий информация может передаваться из межклеточного 713 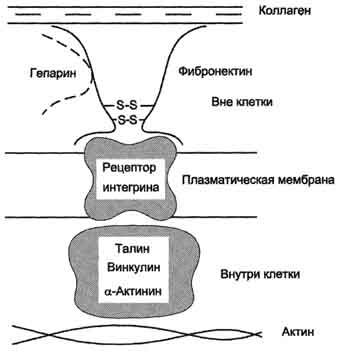 Рис. 15-20. Схема взаимодействия фибронектина с интегрином. матрикса внутрь клетки, а также в обратном направлении - из клетки наружу, таким образом влияя на протекающие в клетке процессы. Известно также, что фибронектин участвует в миграции клеток, которые могут присоединяться к его RGD-участкам, и, таким образом, фибронектин как бы помогает им перемещаться в межклеточном матриксе. В межклеточном матриксе, окружающем трансформированные (или опухолевые) клетки, количество фибронектина заметно снижено, что может быть одной из причин появления метастазов. Ламинин - наиболее распространённый некол-лагеновый гликопротеин базальных мембран. Он состоит из трёх полипетидных цепей: А, В1 и В2. Молекула ламинина имеет крестообразную форму с тремя одноцепочечными ветвями и одной трёхцепочечной ветвью (рис. 15-21). Каждая цепь ламинина содержит несколько глобулярных и стержневидных доменов, на которых имеются специфические центры связывания для различных веществ. Ламинин взаимодействует со всеми структурными компонентами базальных мембран, включая коллаген IV типа, нидоген, фибронектин, ГСПГ. Кроме того, молекула ламинина имеет несколько центров связывания с клетками. Главные функции ламинина определяются его способностью связывать клетки и модулировать клеточное поведение. Он может влиять на рост, морфологию, дифференцировку и подвижность клеток. Ламинин выполняет роль адгезивного белка для различных эпителиальных и мезенхимальных клеток. Гликозаминогликаны- линейные отрицательно заряженные гетерополисахариды. РаНbше их называли мукополисахаридами, так как они обнаруживались в слизистых секретах (мукоза) и придавали этим секретам вязкие, смазочные свойства. Эти свойства обусловлены тем, что гликозаминогликаны могут связывать большие количества воды, в результате чего межклеточное вещество приобретает желеобразный характер. Протеогликаны- высокомолекулярные соединения, состоящие из белка (5-10%) и гликозаминогликанов (90-95%). Они образуют основное вещество межклеточного матрикса соединительной ткани и могут составлять до 30% сухой массы ткани. Белки в протеогликанах представлены одной полипептидной цепью разной молекулярной массы. Полисахаридные компоненты у разных протеогликанов разные. Протеогликаны отличаются от большой группы белков, которые называют гликопротеинами.Эти белки тоже содержат олигосахаридные цепи разной длины, ковалентно 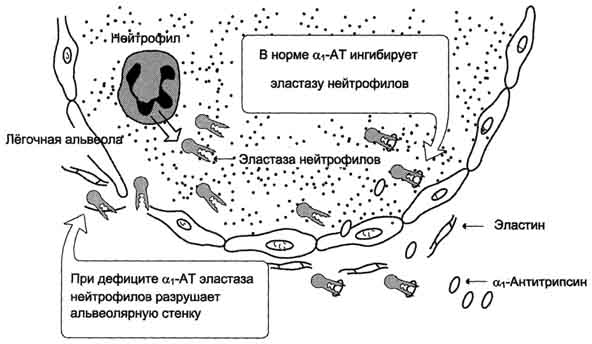 Рис. 15-13. Разрушение лёгочных альвеол эластазой нейтрофилов. 703 присоединённые к полипептидной основе. Углеводный компонент гликопротеинов гораздо меньше по массе, чем у протеогликанов, и составляет не более 40% от общей массы. Гликопротеины выполняют в организме человека разные функции и присутствуют во всех классах белков - ферментах, гормонах, транспортных, структурных белках и др. Представители гликопротеинов - коллаген и эластин, иммуноглобулины, ангиотензиноген, трансферрин, церулоплазмин, внутренний фактор Касла, тиреотропный гормон. Гликозаминогликаны и протеогликаны, являясь обязательными компонентами межклеточного матрикса, играют важную роль в межклеточных взаимодействиях, формировании и поддержании формы клеток и органов, образовании каркаса при формировании тканей. Благодаря особенностям своей структуры и физико-химическим свойствам, протеогликаны и гликозаминогликаны могут выполнять в организме человека следующие функции:
А. Строение и классы гликозаминогликатов Гликозаминогликаны представляют собой длинные неразветвлённые цепи гетерополиса-харидов. Они построены из повторяющихся дисахаридных единиц. Одним мономером этого дисахарида является гексуроновая кислота (D-глюкуроновая кислота или L-идуроновая), вторым мономером - производное аминосахара (глюкоз- или галактозамина). NH2-rpynna аминосахаров обычно ацетилирована, что приводит к исчезновению присущего им положительного заряда. Кроме гиалуроновой кислоты, все гликозаминогликаны содержат сульфатные группы в виде О-эфиров или N-сульфата. В настоящее время известна структура шести основных классов гликозаминогликанов, которые представлены в табл. 15-4. Гиалуроновая кислотанаходится во многих органах и тканях. В хряще она связана с белком и участвует в образовании протеогликановых агрегатов, в некоторых органах (стекловидное тело глаза, пупочный канатик, суставная жидкость) встречается и в свободном виде. Предполагается, что в суставной жидкости гиалуроновая кислота выполняет роль смазочного вещества, уменьшая трение между суставными поверхностями. Повторяющаяся дисахаридная единица в гиалуроновой кислоте имеет следующую структуру: 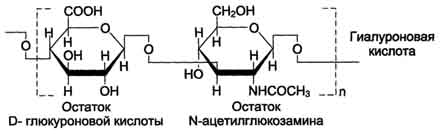 704 Таблица 15-4. Структура различных классов гликозаминогликанов
1 В состав дисахаридной единицы может входить D-глюкуроновая кислота. 2 Может содержать N-сульфопроизводное глюкозамина вместо N-ацетилглюкозамина и различное количество идуроновой и глюкуроновой кислот. 705 Гиалуроновая кислота содержит несколько тысяч дисахаридных единиц, молекулярная масса её достигает 105 - 107 Д. Хондроитинсульфаты- самые распространённые гликозаминогликаны в организме человека; они содержатся в хряще, коже, сухожилиях, связках, артериях, роговице глаза. Хондроитинсульфаты являются важным составным компонентом агрекана - основного протеогликана хрящевого матрикса. В организме человека встречаются 2 вида хондроитинсульфатов: хондроитин-4-сульфат и хондроитин-6-сульфат. Они построены одинаковым образом, отличие касается только положения сульфатной группы в молекуле N-ацетилгалактозамина (см. схему А). Одна полисахаридная цепь хондроитинсульфата содержит около 40 повторяющихся дисахаридных единиц и имеет молекулярную массу 104 - 106 Д. Кератансульфагы- наиболее гетерогенные гликозаминогликаны; отличаются друг от друга по суммарному содержанию углеводов и распределению в разных тканях. Кератансульфат I находится в роговице глаза и содержит кроме повторяющейся дисахаридной единицы L-фукозу, D-маннозу и сиаловую кислоту. Кератансульфат II был обнаружен в хрящевой ткани, костях, межпозвоночных дисках. В его состав помимо Сахаров дисахаридной единицы входят N-ацетилгалактозамин, L-фукоза, D-манноза и сиаловая кислота. Кератансульфат II входит в состав агрекана и некоторых малых протеогликанов хрящевого матрикса. В отличие от других гликозаминогликанов, кератансульфаты вместо гексуроновой кислоты содержат остаток галактозы (см. схему Б). Молекулярная масса одной цепи кератансуль-фата колеблется от 4 × 103 до 20 × 103 Д. Дерматансульфатшироко распространён в тканях животных, особенно он характерен для кожи, кровеносных сосудов, сердечных клапанов. В составе малых протеогликанов (бигликана и декорина) дерматансульфат содержится в межклеточном 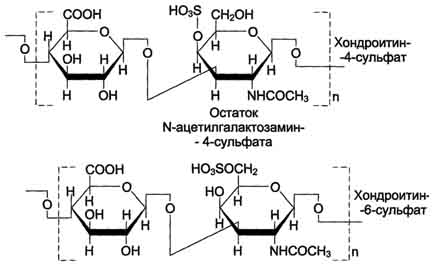 Схема А 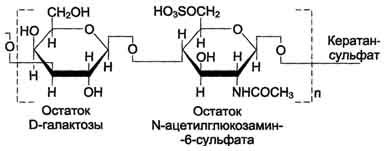 Схема Б 706 веществе хрящей, межпозвоночных дисков, менисков. Повторяющаяся дисахаридная единица дерматансульфата имеет следующую структуру (см. схему А). Молекулярная масса одной цепи дерматансульфата колеблется от 15 × 103 до 40 × 103 Д. Гепарин- важный компонент противосвёртывающей системы крови (его применяют как антикоагулянт при лечении тромбозов). Он синтезируется тучными клетками и находится в гранулах внутри этих клеток. Наибольшие количества гепарина обнаруживаются в лёгких, печени и коже. Дисахаридная единица гепарина похожа на дисахаридную единицу гепарансульфата. Отличие этих гликозаминогликанов заключается в том, что в гепарине больше N-сульфатных групп, а в гепарансульфате больше N-ацетильных групп. Молекулярная масса гепарина колеблется от 6 × 103 до 25 × 103 Д (см. схему Б). Гепарансульфатнаходится во многих органах и тканях. Он входит в состав протеогликанов базальных мембран. Гепарансульфат является постоянным компонентом клеточной поверхности. Структура дисахаридной единицы гепарансульфата такая же, как у гепарина. Молекулярная масса цепи гепарансульфата колеблется от 5 × 103 до 12 × 103 Д. 101. Молекулярная структура миофибрилл. Структура и функция основных белков миофибрилл миозина, актина, тропомиозина, тропонина. В мышечной клетке имеются миофибриллы — особым образом организованные пучки белков, располагающиеся вдоль клетки. Миофибриллы, в свою очередь, построены из белковых нитей (филаментов) двух типов — толстых и тонких. Основным белком толстых нитей является миозин, а тонких — актин. Миозиновые и актиновые нити — главный компонент всех сократительных систем. Электронно-микроскопическое изучение поперечных и продольных срезов мышц обнаружило строго упорядоченное расположение миоэиновых и актиновых нитей в миофибрилле. Функциональной единицей миофибриллы является саркомер — участок миофибриллы (длиной 2,5 мкм) между двумя Z-пластинками. Саркомер включает пучок миозиновых нитей, серединой прикрепленных к М-пластинке (М-линия), и пучки актиновых нитей, прикрепленных к Z-пластинкам. Многие сотни саркомеров образуют миофибриллу. Чередование в миофибрилле участков, содержащих толстые нити, с участками, содержащими тонкие нити (A-диски и I-диски), создает поперечную полосатость мышц. Саркоплазма, вмещающая миофибриллы, пронизана между ними сетью цистерн и трубочек эндоплазматического (саркоплазматического) ретикулума, а также системой поперечных трубочек, которые тесно контактируют с саркоплазматическим ретикулумом, но не сообщаются с ним. СТРОЕНИЕ МИОЗИНОВЫХ НИТЕИ Миозиновые нити образованы белком миозином. Миозин составляет почти половину всех белков скелетной мышцы. Молекула миозина содержит две идентичные тяжелые полипептидные цепи (молекулярная масса каждой 200 ООО) и четыре легкие цепи (молекулярная масса около 20 ООО). Каждая тяжелая цепь на большой части длины с С-конца имеет конформацию а-спирали, и обе спирали скручены друг с другом; эта часть молекулы имеет форму палочки. Противоположные концы каждой цепи (N-концы) имеют глобулярную форму, образуя ≪головки≫ молекулы. К каждой из головок нековалентно присоединены по две легкие цепи. Миозин катализирует гидролиз АТФ; это было установлено Энгельгардтом и Любимовой в 1939 г. Энергия гидролиза используется для сокращения мышцы. Значительно позднее выяснилось, что каталитический активный центр локализован в головках молекулы миозина. Открытие АТФазной активности миозина в высокой степени стимулировало исследования мышечного сокращения, поскольку было первым прямым указанием на источник энергии для сокращения и на роль миозина в использовании этой энергии. Палочкообразные хвосты молекул миозина могут соединяться друг с другом продольно, образуя пучки; головки выступают на поверхности пучка, выстраиваясь вокруг него по спирали. В области М-линии пучки стыкуются ≪хвост к хвосту≫. Так получаются миозиновые нити саркомера, каждая из которых содержит около 400 молекул миозина. СТРОЕНИЕ АКТИНОВЫХ НИТЕЙ В состав актиновых нитей входят белки актин, тропомиозин и тропонин. Основу нитей составляют молекулы актина. Актин — это глобулярный белок с молекулярной массой 43 ООО и шарообразными молекулами диаметром около 5 нм; такая форма актина называется G-актин (глобулярный актин). G-актин содержится и во многих немышечных клетках. Молекулы G-актина могут нековалентно соединяться, образуя фибриллярный актин — F-актин. Форма молекул F-актина напоминает две нитки бус, скрученные друг с другом. В мышечных клетках весь актин находится в форме F-актина. К F-актину могут присоединяться головки миозина, причем на каждой молекуле G-актина в F-актине есть центр связывания миозина. В результате такого взаимодействия в сотни раз увеличивается АТФазная активность миозина. Соединение F-актина с миозином называют актомиозином. Образование связей между миозиновыми и актиновыми нитями в саркомере имеет важное значение в процессе сокращения мышцы. Молекулы другого белка актиновых нитей — тропомиозина — имеют форму палочек длиной 40 нм. Они располагаются вблизи желобков спиральной ленты F-актина, вдоль нее, причем каждая молекула тропомиозина соединена с семью молекулам G-актина, а концами примыкает к соседним молекулам тропомиозина. Третий белок актиновых нитей — тропонин — имеет глобулярную форму; он построен из трех разных субъединиц. Тропонин нековалентно связан с тропомиозином и с актином; на каждую молекулу тропомиозина приходится одна молекула тропонина. Одна из субъединиц тропонина содержит Са-связывающие центры: эта субъединица по строению сходна с кальмодулином. Тонкие нити прикреплены к Z-пластинкам, которые тоже представляют собой белковые структуры. Содержание миозина, актина, тропомиозина и тропонина в миофибриллах равно примерно 55, 25, 15 и 5 % соответственно. 102. Биохимические механизмы мышечного сокращения и расслабления. Роль ионов кальция и других ионов в регуляции мышечного сокращения. Особенности энергетического обмена в мышцах; роль креатинфосфата. Механизм сокращения поперечнополосатой мышечной ткани 1. Разряд мотонейрона. До сокращения мышцы концентрация ионов кальция в саркоплазме составляет 10-8 –10-7 моль/л. 2. Высвобождение нейромедиатора : из нервных окончаний ® ацетилхолин 3.Связывание АХ : связывается с холинорецепторами = никотинрецепторами (одновременно являются Nа-каналами, которые открываются) ® в цитоплазму ток ионов Nа (по градиенту концентрации и градиенту заряда)® потенциал ПМ от -80 мВ до +40 мВ =деполяризация мембраны : а)-активация потенциалзависимых Nа-каналов-®---потенциал действия по трубочкам Т-системы ---мышечное сокращение; б) активация потенциалчувствительных К-каналов (ионы К+ наружу) и потенциал покоя восстанавливается. 4. Генерация потенциала действия в мышечном волокне. Меняют конформацию медленные потенциалчувствительные Са-каналы ПМ = дигидропиридинчувствительные (a1-субъединица) ®Са каналы СР (=рианодиновые RyR): 4 п/п цепи с М.м.> 2 200 кДа, активаторы: низкие (микромолярные) концентрации Са2+, АТФ. ЖК, кофеин, полиамины, ионы тяжелых Ме, ингибиторы: рианодин, ионы Mg 2+, Са2+ в мили/М концентрации 5. Распространение деполяризации внутрь волокна по Т-трубочкам. 6. Высвобождение ионов Са из концевых цистерн СР и их диффузия к толстым и тонким филаментам. Передача сигнала от ПМ к мембране СР реализуется в разных типах мышц через химический тип и через конформационный тип передачи сигнала. Мембраны СР содержат Са-каналы и Са-АТФ-азу, которая активируется тем кальцием, что освобождается из ретикулума через Са-каналы и взаимодействует с регуляторными белками (ТнС, кальмодулин). 7. Связывание Са с ТнС, открывающее миозин-связывающие участки актина. Концентрация Са+2 10-6 - 10-5 моль/л необходима и достаточна для образования I комплекса ТнС- Са+2. Связывание Са+2 изменяет пространственную структуру тропонина и комплекс реагирует с ТнI и ТнТ влияя на их взаимодействие с Тм, образуя II комплекс ТнС- 4Са+2- Тм При изменении конформации Тн открываются активные центры F-актина, который взаимодействует с «головками» миозина, образует актомиозин, обладающий АТФ-азной активностью. 8. Образование поперечных связей между актином и миозином и скольжение тонких филаментов относительно толстых, сопровождающееся укорочением волокна. Актомиозин, гидролизуя АТФ, высвобождает энергию для мышечного сокращения, что обеспечивает продвижение актина по миозину. Каждая из головок генерирует маленькое тянущее усилие (несколько пиконьютонов) ® -усилия суммируются – мышца может развивать достаточно большие напряжения. |