|
1 (1). Первичная структура белка. Зависимость свойств и конформации белков от первичной структуры. Примеры полиморфизма белков, гемоглобин а и F, структурные и функциональные отличия
Многие ферменты для проявления каталитической активности нуждаются в присутствии некоторых веществ непептидной природы — кофакторов. Различают две группы кофакторов: ионы металлов (а также некоторые неорганические анионы) и коферменты, которые представляют собой органические вещества.Примерно треть всех известных ферментов содержит ион металла или активируется ионами металла.
Прочность связи металлов с белковой частью фермента колеблется в широких пределах. Некоторые ферменты в процессе их выделения утрачивают ион металла вследствие диссоциации, так что при измерении активности фермента приходится эти ионы добавлять — это ферменты, активируемые металлами. Другие ферменты сохраняют ион металла при очистке — это металло-ферменты (металлопротеины). Деление на эти группы условно, поскольку между крайними формами существует ряд промежуточных форм.
В роли кофактора могут выступать ионы различных металлов (табл. 2.1). Ион металла может участвовать в присоединении субстрата, собственно в катализе, в стабилизации оптимальной конформации молекулы фермента, в стабилизации четвертичной структуры. Активность металлозависимых ферментов после удаления металла либо утрачивается полностью, либо заметно снижается.Коферменты — это органические вещества, как правило, неаминокислотной природы, непосредственно участвующие в катализе в составе фермента
Ингибирование - частичное или полное торможение ферментативной реакции под действием веществ различной химической природы. Вещества, вызывающие ингибирование ферментов, называют ингибиторами.
Ингибирование - частичное или полное торможение ферментативной реакции под действием веществ различной химической природы. Вещества, вызывающие ингибирование ферментов, называют ингибиторами.
Различают обратимое и необратимое ингибирование. Если ингибитор вызывает стойкое снижение скорости реакции, то это необратимое ингибирование. При этом образуются ковалентные связи между молекулами фермента и ингибитора. Некоторые ферменты полностью ингибируются очень малыми концентрациями ионов тяжёлых металлов, например, ионов ртути (Hg2+), серебра (Ag+) и мышьяка (As+), или иодуксусной кислотой. Эти ингибиторы необратимо соединяются с SH-группами ферментов и вызывают денатурацию ферментного белка.
Диизопропилфторфосфат (ДФФ) – соединение из группы нервнопаралитических отравляющих веществ. Он является ингибитором ацетилхолинэстеразы, которая инактивирует нейромедиатор ацетилхолин. ДФФ связывается с остатком аминокислоты серина в активном центре и блокирует действие фермента. В результате ацетилхолин накапливается в синаптической щели, нервные импульсы следуют один за другим, мышца не расслабляется, и наступает паралич или смерть.
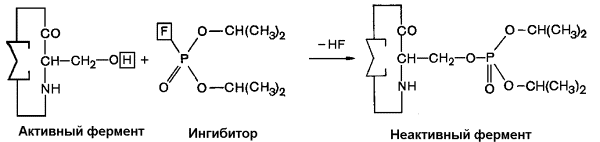
Рисунок 3.16. Необратимое ингибирование фермента ацетилхолинэстеразы диизопрропилфторфосфатом.
Другим примером необратимого ингибирования может служить действие цианидов на фермент цитохромоксидазу, участвующую в окислительно-восстановительных процессах в митохондриях клеток. Отравление цианидами может привести к смерти.
Если ингибитор соединяется с ферментом при помощи нековалентных связей, то возможно восстановление исходной активности фермента после удаления ингибитора, например, путём диализа. Такое ингибирование называется обратимым.
Обратимое ингибирование можно разделить на конкурентное и неконкурентное.
Запомните особенности, характерные для конкурентного ингибирования:
конкурентный ингибитор сходен по строению с субстратом.
конкурентный ингибитор взаимодействует с активным центром фермента, образуя фермент-ингибиторный комплекс, и препятствует взаимодействию активного центра с субстратом.
действие конкурентного ингибитора зависит от его концентрации: чем выше концентрация ингибитора, тем ниже скорость ферментативной реакции.
действие конкурентного ингибитора можно снять, увеличив концентрацию субстрата.
График зависимости скорости ферментативной реакции от концентрации субстрата в присутствии конкурентного ингибитора даёт такую же величину Vmax, как и в отсутствии ингибитора. Величина KM в данном случае будет увеличена, поскольку для обеспечения скорости, равной половине максимальной, в присутствии ингибитора потребуется больше субстрата. Отсюда следует, что конкурентный ингибитор препятствует образованию фермент-субстратного комплекса, но не влияет на процесс распада фермент-субстратного комплекса с образованием продуктов реакции.
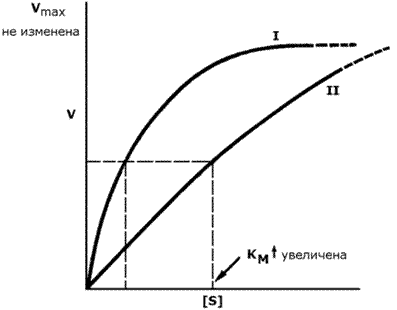
Рисунок 3.17. Влияние конкурентного ингибитора на кинетические свойства фермента.
Примером конкурентного ингибирования является ингибирование фермента сукцинатдегидрогеназы малоновой кислотой. Сукцинатдегидрогеназа катализирует реакцию дегидрирования янтарной кислоты с образованием фумаровой кислоты. Малоновая кислота, как и янтарная кислота, содержит две карбоксильные группы, но обладает более короткой углеродной цепью. Поэтому дегидрирование малоновой кислоты невозможно. Если концентрация малоновой кислоты в среде будет превышать концентрацию янтарной, то активность сукцинатдегидрогеназы снижается. Ингибирующее действие малоновой кислоты исчезает при увеличении концентрации янтарной кислоты.
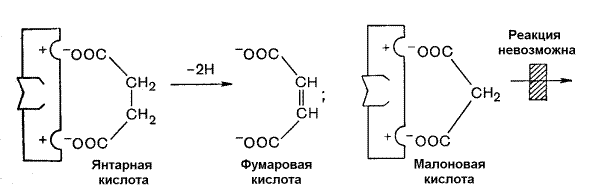
Рисунок 3.18. Конкурентное ингибирование сукцинатдегидрогеназы малоновой кислотой.
Запомните особенности, характерные для неконкурентного ингибирования:
неконкурентный ингибитор не сходен по строению с субстратом.
неконкурентный ингибитор может взаимодействовать, как правило, не с активным центром фермента, а с другими участками в молекуле фермента. Поэтому фермент-ингибиторный комплекс может присоединять субстрат. На ввиду изменения конформации активного центра сродство к субстрату будет понижено.
действие неконкурентного ингибитора не зависит от его концентрации.
действие неконкурентного ингибитора нельзя снять, увеличив концентрацию субстрата.
График зависимости скорости реакции от концентрации субстрата в присутствии неконкурентного ингибитора показывает сниженную величину Vmax. Субстрат не может вытеснить ингибитор из его соединения с ферментом. Величина KM в присутствии неконкурентного ингибитора не меняется. Это значит, что неконкурентный ингибитор воздействует на фермент на стадии распада фермент-субстратного комплекса, но не влияет на связывание субстрата.
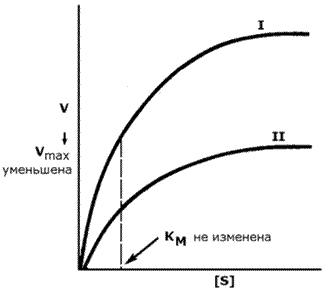
Рисунок 3.19. Влияние неконкурентного ингибитора на кинетические свойства фермента.
Неконкурентные ингибиторы снижают количество молекул субстрата, которые взаимодействуют с одной молекулой фермента в единицу времени (число оборотов фермента).
Ингибиторы ряда ферментов используются в медицине как химиотерапевтические препараты. Целью химиотерапии является уничтожение возбудителя болезни при помощи химических веществ, не повреждая при этом организма-хозяина.
|
5.Применение ингибиторов ферментов
Изучение ингибиторов ферментов - это важная отрасль знания, имеющая фундаментальное значение для фармакологии и токсикологии.
В медицинской практике при лечении многих заболеваний, связанных с нарушениями работы различных ферментов, широко применяются лекарственные препараты, содержащие очень малые дозы ингибиторов соответствующих ферментов. Также широко используются при лечении инфекционных заболеваний сульфаниламидные препараты, обладающие антимикробным действием. Ингибиторы ангиотензинпревращающего фермента широко вошли в клиническую практику с середины 80-х годов, в первую очередь, как препараты с выраженным анти гипертензивным действием, применяемые в лечении сердечно-сосудистых заболеваний (Бочков В.Н., А.Б. Добровольский, 2005).
Оно находит непосредственное практическое применение, в том числе в военном деле: "нервные газы" (например зоман, зарин и др.), представляют собой по существу специфические ингибиторы ферментов.
Использование некоторых ингибиторов ферментов в качестве инсектицидов послужило основой для создания крупной развивающейся отрасли промышленности, и широко применяются в различных областях сельского хозяйства, а также в бытовых условиях.
Ингибиторы нашли широкое применение в энзимологии при исследовании природы множественных форм ферментов и изоферментов.
Изучение действия токсических веществ in vitro, представленных ингибиторами ферментов, оказывается полезным при разработке противоядий (антидотов) к этим ядовитым веществам.
8 (1). Свойства ферментов. Зависимость скорости ферментативной реакции от концентрации фермента и субстрата, температуры и рН среды
Свойства ферментов. Ферменты сочетают в себе свойства катализаторов и свойства, присущие всем остальным белкам.
Как и все белки, ферменты:
состоят из многих сотен аминокислотных остатков, соединённых в определённой последовательности при помощи пептидных связей;
они имеют глобулярную пространственную структуру, определяющую их функциональные возможности;
различаются по молекулярной массе, электрофоретической подвижности,
образуют коллоидные растворы.
Подобно неорганическим катализаторам, ферменты:
не расходуются в процессе реакции,
увеличивают скорость как прямой, так и обратной реакции,
не изменяют положения равновесия.
Белковая природа ферментов обусловливает появление у них ряда свойств, в целом нехарактерных для неорганических катализаторов: олигодинамичность, специфичность, зависимость скорости реакции от температуры, рН среды, концентрации фермента и субстрата, присутствия активаторов и ингибиторов.
Олигодинамичность
Ферменты обладают высокой эффективностью даже в очень небольших количествах. Такая высокая эффективность объясняется тем, что молекулы ферментов в процессе своей каталитической деятельности непрерывно регенерируют. Типичная молекула фермента может регенерировать миллионы раз в минуту. Надо сказать, что и неорганические катализаторы также способны ускорять превращение такого количества веществ, которое во много раз превышает их собственную массу. Но ни один неорганический катализатор не может сравниться с ферментами по эффективности действия.
Примером может служить фермент реннин, вырабатываемый слизистой оболочкой желудка жвачных животных. Одна молекула его за 10 минут при 37°С способна вызывать коагуляцию (створаживание) порядка миллиона молекул казеиногена молока.
Другой пример высокой эффективности ферментов даёт каталаза. Одна молекула этого фермента при 0°С расщепляет за секунду около 50 000 молекул пероксида водорода:
2 Н2О2  2 Н2О + О2 2 Н2О + О2
Действие каталазы на пероксид водорода заключается в изменении величины энергии активации этой реакции приблизительно от 75 кДж/моль без катализатора до 21 кДж/моль в присутствии фермента. Если же в качестве катализатора этой реакции используется коллоидная платина, то энергия активации составляет всего 50 кДж/моль.
При изучении влияния какого-либо фактора на скорость ферментативной реакции все прочие факторы должны оставаться неизменными и по возможности иметь оптимальное значение.
Мерой скорости ферментативных реакций служит количество субстрата, подвергшегося превращению в единицу времени, или количество образовавшегося продукта. Изменение скорости проводят на начальной стадии реакции, когда продукт ещё практически отсутствует, и обратная реакция не идёт. Кроме того, на начальной стадии реакции концентрация субстрата соответствует его исходному количеству.
Зависимость скорости ферментативной реакции (V) от концентрации фермента [Е] (рисунок 2.3). При высокой концентрации субстрата (многократно превышающей концентрацию фермента) и при постоянстве других факторовскорость ферментативной реакции пропорциональна концентрации фермента. Поэтому зная скорость реакции, катализируемой ферментом, можно сделать вывод о его количестве в исследуемом материале.
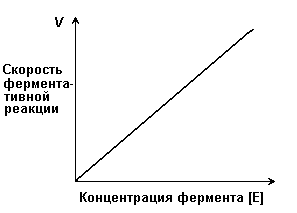
Рисунок 2.3. Зависимость скорости ферментативной реакции от концентрации фермента
Зависимость скорости реакции от концентрации субстрата [S]. График зависимости имеет вид гиперболы (рисунок 2.4). При постоянной концентрации фермента скорость катализируемой реакции возрастает с увеличением концентрации субстрата до максимальной величины Vmax, после чего остаётся постоянной. Это следует объяснить тем, что при высоких концентрациях субстрата все активные центры молекул фермента оказываются связанными с молекулами субстрата. Любое избыточное количество субстрата может соединиться с ферментом лишь после того, как образуется продукт реакции и освободится активный центр.
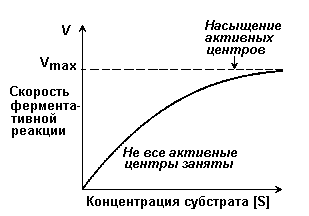
Рисунок 2.4. Зависимость скорости ферментативной реакции от концентрации субстрата.
Зависимость скорости реакции от концентрации субстрата может быть выражена уравнением Михаэлиса — Ментен:
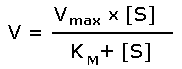 , ,
где V — скорость реакции при концентрации субстрата [S] , Vmax —максимальная скорость и KM —константа Михаэлиса.
Константа Михаэлиса равна концентрации субстрата, при которой скорость реакции составляет половину максимальной. Определение KM и Vmax имеет важное практическое значение, так как позволяет количественно описать большинство ферментативных реакций, включая реакции с участием двух и более субстратов. Различные химические вещества, изменяющие активность ферментов, по-разному воздействуют на величины Vmax и KM.
Зависимость скорости реакции от t – температуры, при которой протекает реакция (рисунок 2.5), имеет сложный характер. Значение температуры, при котором скорость реакции максимальна, представляет собой температурный оптимум фермента. Температурный оптимум большинства ферментов организма человека приблизительно равен 40°С. Для большинства ферментов оптимальная температура равна или выше тойц температуры, при которой находятся клетки.
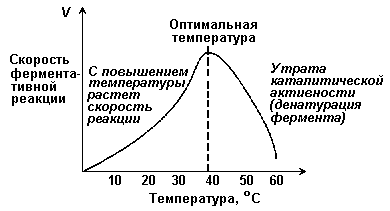
Рисунок 2.5. Зависимость скорости ферментативной реакции от температуры.
При более низких температурах (0° — 40°С) скорость реакции увеличивается с ростом температуры. При повышении температуры на 10°С скорость ферментативной реакции удваивается (температурный коэффициент Q10 равен 2). Повышение скорости реакции объясняется увеличением кинетической энергии молекул. При дальнейшем повышении температуры происходит разрыв связей, поддерживающих вторичную и третичную структуру фермента, то есть тепловая денатурация. Это сопровождается постепенной потерей каталитической активности.
Зависимость скорости реакции от рН среды (рисунок 2.6). При постоянной температуре фермент работает наиболее эффективно в узком интервале рН. Значение рН, при котором скорость реакции максимальна, представляет собой оптимум рН фермента. У большинства ферментов организма человека оптимум рН находится в пределах рН 6 – 8, но есть ферменты, которые активны при значениях рН, лежащих за пределами этого интервала.
Изменение рН как в кислую, так и в щелочную сторону от оптимума приводит к изменению степени ионизации кислых и основных групп аминокислот, входящих в состав фермента (например, СООН-группы аспартата и глутамата, NН2-группы лизина и т.д.). Это вызывает изменение конформации фермента, в результате чего изменяется пространственная структура активного центра и снижение его сродства к субстрату. Кроме того, при экстремальных значениях рН происходит денатурация фермента и его инактивация.
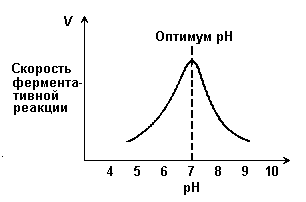
Рисунок 2.6. Зависимость скорости ферментативной реакции от рН среды.
Следует отметить, что свойственный ферменту оптимум рН не всегда совпадает с рН его непосредственного внутриклеточного окружения. Это позволяет предположить, что среда, в которой находится фермент, в какой-то мере регулирует его активность.
Зависимость скорости реакции от присутствия активаторов и ингибиторов. Активаторы повышают скорость ферментативной реакции. Ингибиторы понижают скорость ферментативной реакции.
В качестве активаторов ферментов могут выступать неорганические ионы. Предполагают, что эти ионы заставляют молекулы фермента или субстрата принять конформацию, способствующую образованию фермент-субстратного комплекса. Тем самым увеличивается вероятность взаимодействия фермента и субстрата, а следовательно и скорость реакции, катализируемой ферментом. Так, например, активность амилазы слюны повышается в присутствии хлорид-ионов.
|
9 (1). Основные механизмы регуляции действия ферментов и их роль в регуляции метаболизма. Проферменты
Механизмы действия ферментов
Первоначальным событием при действии фермента является его специфическое связывание с лигандом — субстратом (S). Это происходит в области активного центра, который формируется из нескольких специфических R—групп аминокислот, определенным образом ориентированных в пространстве.
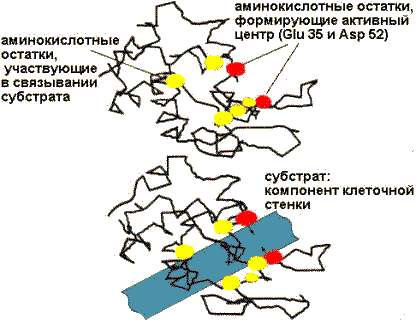
Важнейшие аминокислотные остатки в активном центре лизоцима
|
У некоторых ферментов в активном центре располагается и кофактор. Одни R—группы активного центра принимают участие в связывании субстрата, другие — в катализе. Некоторые группы могут делать и то, и другое. Детальный механизм действия каждого фермента уникален, но есть общие черты в Lработе¦ ферментов, которые заключаются в следующем: высокая избирательность действия фермента обеспечивается тем, что субстрат связывается в активном центре фермента в нескольких точках и это исключает ошибки; активный центр располагается в углублении (нише) поверхности фермента и имеет комплементарную субстрату конфигурацию. В результате субстрат окружается функциональными группами активного центра фермента и изолируется от водной среды. Связывание субстрата с ферментом часто вызывает конформационные изменения, что ведет к правильному (оптимальному) расположению аминокислотных остатков, требуемому для протекания катализа, и тем самым увеличивает специфичность фермент—субстратного взаимодействия (индуцированное соответствие). Так как максимальная активность фермента обусловлена оптимальной конформацией молекулы фермента в целом и активного центра в частности, то даже небольшие изменения окружающих условий, которые затрагивают связывание субстрата или конформацию третичной структуры белка, будут влиять на скорость ферментативной реакции. Например, изменение рН приводит к изменению степени ионизации ионогенных групп фермента и, следовательно, ведет к перераспределению межрадикальных связей в третичной структуре. Оптимальное рН для каждого фермента означает некоторое оптимальное состояние его ионизации, соответствующее наилучшей комплементарности. Изменение температуры вызывает противоречивый эффект: с одной стороны, при повышении температуры до 37 — 40о скорость ферментативной реакции увеличивается, что закономерно для катализа; с другой стороны, при температуре более 50о начинается денатурация фермента.
.
Проферменты, или проэнзимы, зимогены, энзимогены — функционально неактивные предшественники ферментов, подвергающиеся тем или иным преобразованиям (обычно расщеплению специфическими эндо- или экзопептидазами или гидролизу), в результате чего образуется каталитически активный продукт — фермент. Относятся к группе протеиназ(сериновые, тиоловые, кислые). Синтез зимогенов осуществляется на рибосомах эндоплазматического ретикулума особыми секреторными клетками в виде зимогенных гранул, которые после завершения процесса мигрируют к поверхности клеток и затем секретируются в окружающую среду. Достигнув места действия они превращаются в активные формы ферментов. К ним относятся пепсиноген, активной формой которого является пепсин (основной протеолитический фермент желудочного сока), трипсиноген — трипсин, химотрипсиноген — химотрипсин, прокарбоксилепептидазы — карбоксипептидазы (ферменты поджелудочной железы) и др. К зимогенам относятся ферменты свёртывания крови (факторы свёртывания крови), компоненты и факторы системы комплемента
10 (2). Принципы количественного определения ферментов. Единицы ак�тивности ферментов. Основные направления использования ферментов в медицине. Эн�зимодиагностика, энзимотерапия, использование ферментов как реагентов.
Принципы количественного определения активности ферментов. Единицы измерения активности ферментов(1 катал = 1 моль субстрата, превращенного за 1 секунду; 1 международная единица = 1 микромоль субстрата, превращенного за 1 минуту).
Общие методы определения активности ферментов.
Прежде чем преступить к выделению фермента, необходимо избрать и тщательно отработать метод определения активности, под контролем которого производится выбор наиболее эффективных приемов очистки ферментов, а затем и выполнение последовательных стадий его препаративного получения. Активность фермента меняется при различных условиях реакции и зависит от температуры, рН среды, от концентраций субстратов и кофакторов. Учитывая это, при определении активности фермента на разных стадиях очистки необходимо строго соблюдать одни и те же условия. Желательно не ограничиваться определением активности по одному какому-либо методу. Количество субстрата, превращаемого в условиях теста по определению активности фермента, должно быть пропорционально количеству последнего и времени инкубирования. Если же нет такой пропорциональности, то активность рассчитывают по предварительно построенному калибровочному графику, отражающему зависимость скорости реакции от количества единиц фермента. Когда ход реакции нелинеен во времени, следует определять начальную скорость реакции (по тангенсу угла наклона касательной к начальному отрезку кривой превращения). Для этого легче всего применять такие методы изменения активности, которые позволяют непрерывно следить за ходом превращения во времени: спектрофотометрические методы, потенциометрические, полярографические и т.п. Для измерения скорости ферментативной реакции необходимо выбрать буфер, который не тормозит исследуемую активность и обеспечивает поддержание рН раствора, близкой к оптимальной для данного фермента. Реакцию проводят при температуре, лежащей в большинстве случаев в пределах 25-400С. При исследовании ферментов, требующих присутствия кофакторов (ионов металлов, коферментов и др.), концентрация которых может снижаться по мере очистки фермента, в реакционную смесь следует добавлять недостающие кофакторы, например соли металлов, АТФ, НАДФ и т.п. Также для определения активности ферментов вводят стабилизаторы в состав опытных смесей. Во многих случаях добавление желатина, альбумина и других добавок предотвращает денатурацию ферментного белка.
1. Некоторые ферменты применяют как лечебные препараты:
- при их отсутствии или недостатке (наследственном или приобретенном), например, ферменты пищева�рительного тракта (пепсин, трипсин, липаза) входят в состав лекарств, улучшающих переваривание;
- для специфического разрушения некоторых продуктов обмена (например, мочевины), тром�бов, участков омертвевшей ткани на ранах.
Ферменты используют в клинико-диагностичес�ких лабораторияхдля измерения в крови концент�рации глюкозы, жира, холестерина, активных форм кислорода и других веществ.
Такой анализ занимает несколько минут, при этом используется всего 10 мкл плазмы крови.
Для анализа часто используют иммобилизован�ные ферменты, которые искусственно связаны с нерастворимым в воде носителем, что повышает стабильность белковых катализаторов.
В клинических лабораториях применяют гото�вые наборы реактивов, в состав которых входят ферменты, составляющие буфер соли, а также ко�факторы. При использовании иммобилизованных ферментов для определения концентрации компо�нента крови достаточно нанести образец сыворот�ки на индикаторную пластинку и сравнить интен�сивность окраски с эталоном.
. Существует связь между степенью поврежде�ния клеток и последовательностью выхода в кровь ферментов из разных отделов клетки. При воспалительных процессахповышается прони�цаемость клеточных мембран и в сыворотку крови могут попадать цитоплазматические ферменты.
Некроз(омертвение) ткани сопровождается раз�рушением всех клеточных структур, и в сыворотке могут быть обнаружены митохондриальные, ядер�ные и другие ферменты.
Энзимодиагностика.
Некоторые ферменты, проферменты и их субстраты в норме постоянно циркулируют в крови человека и выполняют физиологические функции. Примерами таких ферментов плазмы являются липопротеинлипаза, псевдохолинэстераза, а также проферменты компонентов систем свертывания крови и растворения кровяного сгустка. Эти ферменты называются секреторными, большинство из них синтезируется в печени и секретируются в кровь. Концентрация таких энзимов в крови либо такая же, как в тканях, либо более высокая.
Кроме секреторных ферментов, в плазме крови присутствуют ферменты, которые не выполняют в крови никаких известных физиологических функций. Их субстраты в плазме обычно не обнаруживаются, и в норме их концентрация в крови человека почти в миллион раз ниже, чем в тканях. Такие ферменты называются индикаторными. Появление их в плазме крови в повышенных концентрациях указывает на повышенную скорость разрушения тканей. Таким образом, измерение в крови уровня индикаторных ферментов плазмы дает врачу ценную диагностическую и прогностическую информацию.
Индикаторные ферменты обычно обнаруживаются в плазме в малых количествах, и появляются в ней вследствие постоянно протекающих процессов разрушения клеток организма. Появление этих ферментов в плазме в повышенных концентрациях указывает на повышенную скорость разрушения тканей. Поступлением в плазму значительных количеств мышечных ферментов сопровождается и выполнение тяжелой физической работы. Для клеток разных органов характерен свой набор ферментов, поэтому повышение в крови активности того или иного фермента может указывать на заболевание соответствующего органа.
В клинической практике используется количественное определение различных индикаторных ферментов плазмы. В клетках поджелудочной железы вырабатываются ферменты трипсин (трипсиноген), липаза и амилаза; при остром воспалении поджелудочной железы (острый панкреатит) активность этих ферментов в крови возрастает; повышенная активность амилазы обнаруживается и в моче.
Довольно часто в целях диагностики проводят анализ изоферментного спектра некоторых ферментов, в частности ЛДГ. В клетках миокарда преобладает изофермент ЛДГ1. При заболеваниях, связанных с повреждением сердечной мышцы, концентрация и активность этого изофермента в плазме крови значительно возрастает. При некоторых заболеваниях печени (в том числе инфекционной природы) в крови возрастает содержание ЛДГ4 и ЛДГ5 - изоферментов, характерных для клеток печени.
В настоящее время для получения этой ценной диагностической и прогностической информации в большинстве случаев используются автоанализаторы. В таблице 4.1 приведен перечень ферментов, активность которых чаще всего исследуют в диагностической энзимологии.
Таблица 4.1.
Основные ферменты сыворотки, используемые в клинической диагностике.
Фермент
|
Заболевание
|
Аспартатаминотрансфераза
|
Инфаркт миокарда
|
Аланинаминотрансфераза
|
Вирусный гепатит
|
Амилаза
|
Острый панкреатит
|
Церулоплазмин
|
Гепатолентикулярная дегенерация (болезнь Вильсона)
|
Креатинфосфокиназа
|
Заболевание мышц и инфаркт миокарда
|
γ-Глутамилтранспептидаза
|
Различные заболевания печени
|
Лактатдегидрогеназа (изозимы)
|
Инфаркт миокарда
|
Липаза
|
Острый панкреатит
|
Кислая фосфатаза
|
Рак предстательной железы
|
Щелочная фосфатаза (изозимы)
|
Различные заболевания костей, закупорка протоков печени
|
Высокая специфичность ферментов позволяет использовать их для обнаружения и количественного определения нормальных и патологических компонентов крови и мочи. Например, с помощью фермента уреазы, действующего только на мочевину, можно проводить определение этого вещества в биологических жидкостях. Фермент глюкозооксидаза применяется для определения глюкозы в крови и моче у больных сахарным диабетом.
ЭНЗИМОТЕРАПИЯ — применение ферментов с леч. целью . При гнойно-некротических процессах (для лечения гнойных ран, трофических язв, пролежней), а также в качестве отхаркивающих средств используют протеолитические ферменты — трипсин, химотрипсин. Для улучшения процессов пищеварения используют препараты, содержащие ферменты желудочно-кишечного тракта, — пепсин, панкреатин, фестал и др. Фибринолитические ферменты (напр., стрептокиназа) используются для растворения тромбов (см. Фибринолитические средства). Лидазу (препарат, содержащий фермент гиалуронидазу расщепляющий один из осн. элементов соединительной ткани — гиалуроновую кислоту) применяют при контрактурах, для рассасывания гематом и рубцов (после травм, ожогов, операций).
11 (1). Катаболические и анаболические пути обмена. Функции метаболизма. Три стадии катабо�лизма основных питательных веществ в организме. Связь общего пути катаболизма с цепью переноса электронов и протонов и синтезом АТФ. Роль НАД- и ФАД-зависимых дегидрогеназ
. Метаболизм складывается из двух противоположных сторон: катаболизма и анаболизма.
Катаболизм – это фаза, в которой происходит последовательное расщепление сложных молекул до более простых, таких, как СО2, вода и аммиак. Процессы катаболизма сопровождаются выделением энергии. Эта энергия частично аккумулируется в форме макроэргического соединения – аденозинтрифосфата (АТФ).
Анаболизм – это фаза метаболизма, в которой происходит образование (биосинтез) сложных молекул (белков, липидов, полисахаридов) из простых предшественников. Процессы биосинтеза протекают с затратой энергии. Источником этой энергии служит распад АТФ до АДФ и неорганического фосфата.
Метаболические пути, выполняющие как катаболическую, так и анаболическую функцию, называют амфиболическими.
8.1.5. Катаболическая и анаболическая фазы метаболизма тесно связаны между собой:
а) Энергия, выделяемая в реакциях катаболизма, и аккумулированная в форме молекул АТФ, потребляется в анаболических процессах.
б) В реакциях катаболизма образуются простые метаболиты, которые могут использоваться в реакциях биосинтеза (анаболизма).
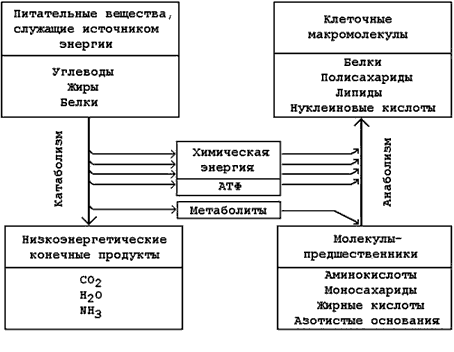
Рисунок 8.2. Взаимосвязь путей катаболизма и анаболизма
. Метаболизм (обмен веществ) – совокупность химических реакций, протекающих в живой клетке. Эти реакции протекают в определённой последовательности и тесно связаны между собой. Главные функции метаболизма в клетке:
а) запасание энергии, которая добывается путем расщепления пищевых веществ, поступающих в организм, или путем преобразования энергии солнечного света;
б) превращение молекул пищевых веществ в строительные блоки;
в) сборку белков, нуклеиновых кислот, липидов, полисахаридов и прочих клеточных компонентов из этих строительных блоков;
г) синтез и разрушение тех биомолекул, которые необходимы для выполнения специфических функций данной клетки
катаболизм – совокупность химических реакций превращения высокомолекулярных соединений в низкомолекулярные. Это сложный ферментативный процесс, в котором принято выделять три основные стадии (рисунок 8.3). Для каждой стадии запомните, какие вещества вступают и образуются, локализацию в организме и энергетическую значимость.
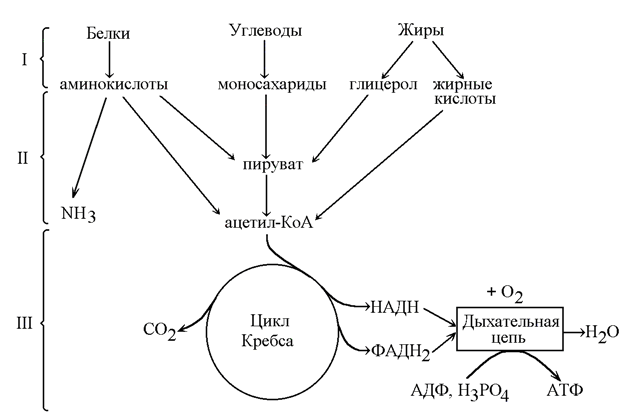
Рисунок 8.3. Три стадии катаболизма питательных веществ (обозначены римскими цифрами).
8.2.2. На первой стадии крупные биомолекулы расщепляются на составляющие их строительные блоки: полисахариды превращаются в пентозы и гексозы, жиры – в жирные кислоты, глицерол и другие компоненты, белки – в аминокислоты. Это происходит в желудочно-кишечном тракте, а также в лизосомах клетки. Реакции катализируют ферменты, относящиеся к классу гидролаз. Относительная энергоотдача составляет менее 1% всей высвобождаемой энергии.
На второй стадии строительные блоки превращаются в более простые молекулы. Моносахариды, глицерол и большинство аминокислот расщепляются до одного и того же трёхуглеродного метаболита – пирувата. Это происходит в цитоплазме клеток. В дальнейшем пируват, а также жирные кислоты и некоторые аминокислоты окисляются до ацетильного остатка, связанного с коэнзимом А (ацетил-КоА). Эти реакции протекают уже в митохондриях клетки. Пируват и ацетил-КоА, находящиеся на пересечении нескольких метаболических путей, можно отнести к ключевым или узловым метаболитам. Относительная энергоотдача второй стадии катаболизма около 20%; выделяемая энергия может быть частично аккумулирована в виде АТФ.
На третьей стадии происходит окисление ацетильной группы в цикле трикарбоновых кислот Кребса до СО2 и восстановленных форм коферментов НАД и ФАД. Эти коферменты окисляются в дыхательной цепи до Н2О; выделяемая энергия аккумулируется в АТФ. Все эти реакции протекают в митохондриях. Относительная энергоотдача третьей стадии - около 80%.
8.2.3. Различают общий и специфические пути катаболизма. К специфическим путям катаболизма веществ того или иного класса относят главным образом реакции первой и второй стадии катаболизма, которые для каждого класса могут существенно различаться. Третья стадия, одинаковая для всех классов питательных веществ, называется общим путём катаболизма.
Тип участвующей дегидрогеназы зависит от природы субстрата. Но каким бы ни был исходный субстрат, электроны и протоны от флавинов переносятся к коферменту Q, а дальше пути электронов и протонов расходятся. Электроны с помощью системы цитохромов достигают кислорода, который затем, присоединяя протоны, превращается в воду. Чтобы разобраться в системе транспорта электронов, необходимо познакомиться с отдельными ее участниками. NAD - зависимая дегидрогеназакатализирует реакции окисления непосредственно субстрата (первичная дегидрогеназа). NAD+ является коферментом и выполняет роль акцептора водорода:
Коферменты дегидрогеназ.Символ 2Н+ означает два электрона и два протона, обычно переносимые в виде гидрид иона. В этом случае вместо терминов «донор электронов» и «акцептор электронов» иногда используют термины «донор или акцептор водорода». FAD - зависимая дегидрогеназа также выполняет функцию первичной дегидрогеназы. Коферментом является FAD, который является акцептором водорода от субстрата. NADH - дегидрогеназа катализирует окисление NADH и восстановление убихинона (CoQ). Переносчиком водорода является кофермент - FMN (комплекс 1). В процессе реакции водород сначала присоединяется к FMN, соединенному с ферментом, а затем передается на убихинон. Флавиновые коферменты (FAD и FMN) прочно связаны с ферментом как простетические группы, поэтому ферменты, в состав которых они входят, называютсяфлавопротеины. Флавинмононуклеотид (FMN), или рибофлавин фосфат, неразрывно связан с белковой частью фермента. Строго говоря, FMN не является нуклеотидом, так как флавиновая часть связана с рибитолом, а не с рибозой.
12 (1). Пировиноградная кислота: пути образования и использования в ор�ганизме. Окислительное декарбоксилирование пировиноградной кислоты. Строение пируватдегидрогеназного комплекса. Значение витаминных ко�ферментов в декарбоксилировании пирувата.
Пировиноградная кислота — химическое соединение с формулой СН3СОСООН, органическая кетокислота.
Пируваты (соли пировиноградной кислоты) — важные химические соединения в биохимии. Они являются конечным продуктом метаболизмаглюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным.
В условиях достаточного поступления кислорода, пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций, известных как цикл Кребса, или дыхательный цикл, цикл трикарбоновых кислот. Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Оксалоацетат затем окисляется до углекислого газа и воды. Эти реакции названы по имени Ханса Адольфа Кребса, биохимика, получившего вместе с Фрицем Липманном Нобелевскую премию по физиологии в 1953 году за исследования биохимических процессов клетки. Цикл Кребса называют также циклом лимонной кислоты, поскольку лимонная кислота является одним из промежуточных продуктов цепи реакций цикла Кребса.
Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты у животных иэтанола[1] у растений.[2] При анаэробном дыхании в клетках пируват, полученный при гликолизе, преобразуется в лактат при помощи ферменталактатдегидрогеназы и NADP в процессе лактатной ферментации, либо в ацетальдегид и затем в этанол в процессе алкогольной ферментации.
Пировиноградная кислота является «точкой пересечения» многих метаболических путей. Пируват может быть превращён обратно в глюкозу в процессе глюконеогенеза, или в жирные кислоты или энергию через ацетил-КоА, в аминокислоту аланин, или в этанол. Таким образом пируват объединяет несколько ключевых метаболических процессов клетки и является универсальной «метаболической валютой» живой клетки.
. Заключительной реакцией второй стадии катаболизма питательных веществ является окислительное декарбоксилирование пирувата.
Пируват подвергается окислительному декарбоксилированию до ацетил-коэнзима А (ацетил-КоА) в митохондриях клеток при участии пируватдегидрогеназного комплекса. Пируватдегидрогеназный комплекс – мультифермент, в состав которого входит 3 фермента:
1) пируватдекарбоксилаза (коферментом служит производное витамина В1 - тиаминдифосфат, ТДФ, формулу его см. в теме «Коферменты»);
2) трансацилаза (кофермент – липоевая кислота, ЛК, см. рисунок 8.4);
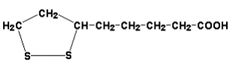
Рисунок 8.4. Липоевая кислота (витаминоподобное вещество).
3) дигидролипоилдегидрогеназа (в качестве кофермента используется производное витамина В2 – флавинадениндинуклеотид, ФАД, формулу его см. в теме «Коферменты»).
В реакции участвуют также два кофермента: коэнзим А (НSКоА, производное витамина В3) и никотинамидадениндинуклеотид (НАД, производное витамина РР), связанные при помощи нековалентных связей соответственно с трансацилазой и дигидролипоилдегидрогеназой.
8.3.2. Суммарное уравнение реакции, катализируемой пируватдегидрогеназным комплексом:
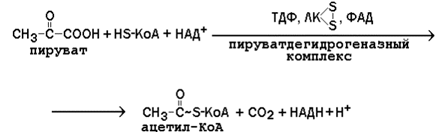
Схема работы этого мультиферментного комплекса представлена на рисунке 8.5.
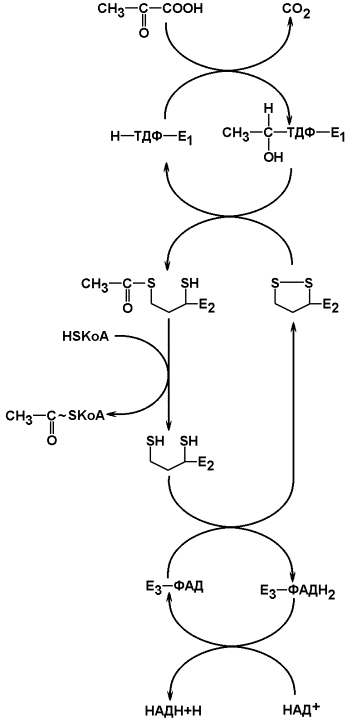
Рисунок 8.5. Окислительное декарбоксилирование пирувата (схема).
Ацетил-КоА затем подвергается окислению в цикле Кребса, а НАДН служит донором водорода для дыхательной цепи. Процесс является аэробным, т.к. конечный акцептор водорода НАДН – кислород. Скорость реакции снижается при накоплении в клетке ацетил-КоА, НАДН и АТФ, увеличивается – при возрастании концентрации АДФ. Регуляторным ферментом комплекса является пируватдекарбоксилаза.
Аналогично происходит окислительное декарбоксилирование α-кетоглутарата – одного из метаболитов цикла Кребса. Реакцию катализирует α-кетоглутаратдегидрогеназный комплекс, сходный с пируватдегидрогеназным комплексом (см. параграф 8.4, рис. 8.7, реакция 5).
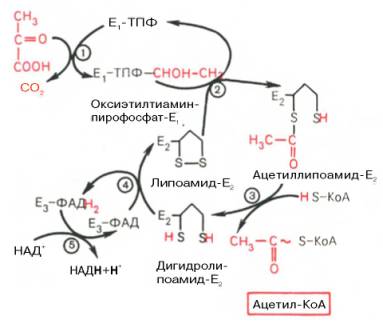
В процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий: в нем принимают участие 3 фермента (пируватдегидрогеназа. Дигидролипоилацетилтрансфераза, дегидролипоилдегидрогеназа) и 5 коферментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД, НАД), из которых три-относительно прочно связаны с ферментами (ТПФ- Е1, липоамид-Е2 и ФАД-Е3), а два- легко диссоциируют (HS-KoA и НАД). Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано,что составляющие комплекс полипептидные цепи субъединиц дигидролипоилацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогенеза.
13 (1). Ацетил-КоА: источники и основные пути использования в тканях. Компартментализация обмена ацетил-КоА.
Ацетил-кофермент А, ацетил-коэнзим А, сокращённо ацетил-КоА — важное соединение в обмене веществ, используемое во многихбиохимических реакциях. Его главная функция — доставлять атомы углерода с ацетил-группой в цикл трикарбоновых кислот, чтобы те былиокислены с выделением энергии. По своей химической структуре ацетил-КоА — тиоэфир между коферментом А (тиолом) и уксусной кислотой(носителем ацильной группы). Ацетил-КоА образуется во время второго шага кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот.
Ацетил-КоА — важный компонент биологического синтеза нейротрансмиттера ацетилхолина. Холин, в соединении с ацетил-КоА, катализируется ферментом холинацетилтрансферазой, чтобы образовать ацетилхолин и коэнзим А.
| |
|
|
 Скачать 1.58 Mb.
Скачать 1.58 Mb.