1 (1). Первичная структура белка. Зависимость свойств и конформации белков от первичной структуры. Примеры полиморфизма белков, гемоглобин а и F, структурные и функциональные отличия
 Скачать 1.58 Mb. Скачать 1.58 Mb.
|

ГалактоземияЭтиология и патогенез.Наследственная энзимопатия. Наследуется по рецессивному типу. В основе галактоземии лежит нарушение обмена галактозы в связи с отсутствием фермента галактозофосфатуридилтрансферазы. В результате в крови накапливается в больших концентрациях галактоза и галактозофосфат. Развивается эндогенная интоксикация, приводящая к поражению головного мозга, печени, хрусталика глаза, почек. Клиническая картина. Клинические признаки заболевания возникают рано, через 1—2 нед после рождения ребенка. Пропадает аппетит, появляются вялость, рвота. Масса тела ребенка уменьшается, ребенок истощается. Постепенно увеличиваются печень, селезенка, может возникнуть желтуха. Часто отмечается катаракта, ведущая к слепоте. Могут быть симптомы, свидетельствующие о поражении почек. Развивается умственная отсталость. При несвоевременной диагностике заболевание прогрессирует, что приводит к тяжелым последствия или летальному исходу. Диагноз. Определение избыточных количеств галактозы в крови и моче. Исследование концентрации галактозофосфатуридилтрансферазы в эритроцитах. Лечение.Диетотерапия является единственным методом лечения. Для вскармливания ребенка используют смеси, лишенные лактозы (см.Алактаэия).Из питания детей более старшего возраста исключают цельномолочные продукты Непереносимость сахарозы Этиология и патогенез. Наследственная болезнь. Встречается чаще врожденной непереносимости лактозы. Наследуете по аутосомно-рецессивному типу. Заболевание связано с дефектом фермента сахаразы (инвертазы) в слизистой оболочке толстой кишки, вследствие чего нарушаете» пристеночное расщепление сахарозы на глюкозу и фруктозу. Клиническая картина. Проявляется с момента введения в питание ребенка молочных смесей, фруктовых соков, содержащих сахарозу, подслащенной воды. Обычно совпадает по времени с началом искусственного вскармливания или прикорма. Основным симптомом является профузная водянистая диарея с кислым запахом, которая осложняется дегидратацией, и повышением температуры тела и коллапсом. Возможно развитие гипотрофии. С возрастом выраженность симптомов уменьшается. Диагноз. Обнаружение сахарозы в каловых массах («Clinitest» после обработки фекалий хлористоводородной кислотой или хроматографический метод), плоская гликемическая кривая и понос после нагрузки сахарозой (2 г/кг), отсутствие активности сахарозы в биоптате тонкой кишки. Лечение. Вскармливание грудным молоком или молочными смесями, имеющими в качестве углеводного компонента исключительно лактозу. Исключение сладких фруктов и соков, сладких блюд. В случае изомальтазной недостаточности — ограничение крахмалсодержащих продуктов и блюд (каши, картофель). Непереносимость лактозы (или гиполактазия) — термин для описания патологических состояний, вызванных снижением уровня лактазы —фермента, необходимого для правильного переваривания лактозы Симптомы лактазной недостаточности определяются избыточным ростом и усилением жизнедеятельности микрофлоры кишечника, усваивающей лактозу, а также осмотическим эффектом непереваренной лактозы в кишечнике (задержка воды в каловых массах). Основные симптомы лактазной недостаточности: метеоризм (вздутие живота), боли в животе,диарея, реже рвота. У детей лактазная недостаточность может проявляться хроническими запорами, беспокойством и плачем после еды. Следует отметить, что симптомы лактазной недостаточности всегда связаны с употреблением в пищу продуктов, содержащих лактозу. 17 (1). Образование и пути использования глюкозо-6-фосфата в организме. Особенности обмена глюкозо-6-фосфата в различных тканях, обусловленные функциональными различиями этих тканей. Глюкозо-6-фосфат образуется в организме разными путями. Во-первых, он может синтезироваться путем фосфорилирования глюкозы за счет ее взаимодействия с АТФ. Во-вторых, он образуется в результате реакцииизомеризации фосфорных эфиров других изомерных ему гексозофосфорных эфиров. В-третьих, он получается из глюкозо-1-фосфата, который представляет собой продукт фосфоролиза олиго- и полисахаридов. Две первые реакции рассмотрены в предыдущем разделе. Что касается преобразования глюкозо-1-фосфата в глюкозо-6-фосфат, то эта реакция протекает в два этапа при участии фермента фосфоглюкомутазы. Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках эпителия кишечника. В клетках этих органов имеется фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём:Глюкозо-6-фосфат +Н2О→Глюкоза + Н3РО4Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку - мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки. Глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основнымииз которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О или лактата, синтез пентоз. Распад глюкозы до конечных продуктов служит источником энергии для организма. Вместе с тем в процессе метаболизма глюкозо-6-фосфата образуются промежуточные продукты, используемые в дальнейшем для синтеза аминокислот, нуклеотидов, глицерина и жирных кислот. Таким образом, глюкозо-6-фосфат - не только субстрат для окисления, но и строительный материал для синтеза новых соединений . 18 (1). Гликолиз - локализация в клетке и тканях, последовательность ре акций, биологическая роль, энергетический баланс. Утилизация молочной кислоты в организме человека. . Гликолиз – это ферментативный распад глюкозы до молочной кислоты (лактата). Гликолиз протекает в тканях без потребления кислорода. В анаэробных условиях гликолиз – единственный процесс, поставляющий АТФ, так как окислительное фосфорилирование в этих условиях не функционирует. Гликолиз протекает в цитоплазме клеток организма. Этот процесс катализируется одиннадцатью ферментами. Условно можно разделить гликолиз на две стадии. 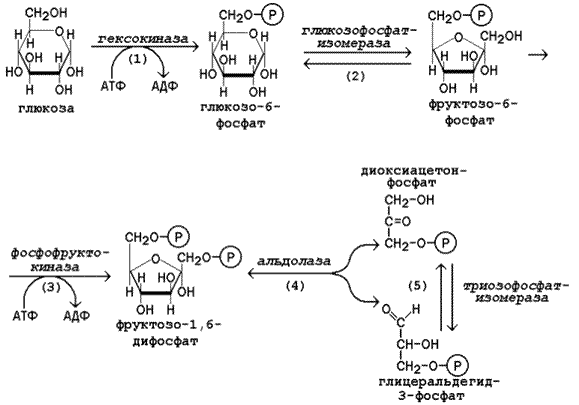 Рисунок 10.3. Реакции первой стадии гликолиза. 10.3.2. Первая стадия гликолиза включает реакции превращения молекулы глюкозы в две молекулы фосфотриоз. Эта стадия сопровождается затратой молекул АТФ. Начальной реакцией глюкозы в клетке является её фосфорилирование в результате взаимодействия с АТФ (рисунок 10.3, реакция 1). Эта реакция в условиях клетки протекает только в одном направлении. Биологическая роль реакции фосфорилирования глюкозы заключается в том, что глюкозо-6-фосфат, в отличие от свободной глюкозы, не может проникать через клеточную мембрану обратно в кровь. В большинстве тканей реакцию фосфорилирования глюкозы катализирует фермент гексокиназа, которая обладает высоким сродством к глюкозе, способна также фосфорилировать фруктозу и маннозу и ингибируется избытком глюкозо-6-фосфата. В клетках печени, кроме того, есть фермент глюкокиназа, которая имеет низкое сродство к глюкозе, не ингибируется глюкозо-6-фосфатом и не участвует в фосфорилировании других моносахаридов. В следующей реакции глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат (рисунок 10.3, реакция 2). Продукт реакции изомеризации подвергается повторному фосфорилированию за счёт АТФ (рисунок 10.3, реакция 3). Эта реакция – наиболее медленно протекающая реакция гликолиза и, подобно фосфорилированию глюкозы, необратима. Фермент – фосфофруктокиназа – является аллостерическим, активируется АДФ и АМФ, ингибируется цитратом и высокой концентрацией АТФ. На следующем этапе фруктозо-1,6-дифосфат подвергается расщеплению на две фосфотриозы (рисунок 10.3, реакция 4). Таким образом, химическое соединение, содержащее 6 углеродных атомов, превращается в два, содержащих по 3 атома углерода. Поэтому гликолиз называют дихотомическим путём превращения глюкозы (от слова дихотомия – рассечение на две части). Далее происходит изомеризация триозофосфатов (рисунок 10.3, реакция 5). В этой реакции диоксиацетонфосфат переходит в глицеральдегид-3-фосфат. Таким образом, в первой стадии гликолиза молекула глюкозы превращается в две молекулы глицеральдегид-3-фосфата. 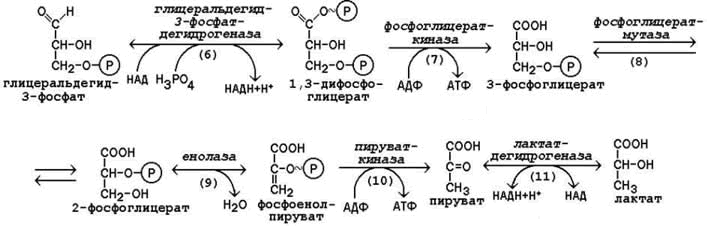 Рисунок 10.4. Реакции второй стадии гликолиза. 10.3.3. Вторая стадия гликолиза включает реакции превращения двух молекул глицеральдегид-3-фосфата в две молекулы лактата. На этой стадии гликолиза происходит синтез молекул АТФ. Глицеральдегид-3-фосфат подвергается дегидрированию при участии НАД-зависимой дегидрогеназы. В этой реакции происходит потребление неорганического фосфата, который включается в состав продукта реакции, содержащего макроэргическую фосфатную связь (рисунок 10.4, реакция 6). 1,3-Дифосфоглицерат вступает в реакцию первого субстратного фосфорилирования, т.е. не сопряжённого с переносом электронов в дыхательной цепи. В этой реакции осуществляется синтез молекулы АТФ в результате переноса фосфатной группы вместе с макроэргической связью на молекулу АДФ (рисунок 10.4, реакция 7). В следующей реакции происходит внутримолекулярное перемещение фосфатной группы 3-фосфоглицерата ко 2-му углеродному атому (рисунок 4, реакция 8). Тем самым облегчается последующее отщепление молекулы воды, которое приводит к появлению в продукте реакции макроэргической фосфатной связи (рисунок 10.4, реакция 9). Фосфоенолпируват (ФЕП) вступает в реакцию второго субстратного фосфорилирования, в ходе которого образуется молекула АТФ. В отличие от первого субстратного фосфорилирования, данная реакция является необратимой в условиях клетки. Фермент пируваткиназа аллостерически ингибируется АТФ и фруктозо-1,6-дифосфатом (рисунок 10.4, реакция 10). В заключительной реакции гликолиза происходит использование НАДН, образовавшегося при дегидрировании глицеральдегид-3-фосфата (см. реакцию 6). При участии НАД-зависимой лактатдегидрогеназы пируват восстанавливается в молочную кислоту (рисунок 10.4, реакция 11). Фермент существует в пяти изоферментных формах, отличающихся сродством к субстрату и распределением в тканях. Таким образом, в процессе гликолиза в клетке не накапливается НАДН. Это значит, что гликолиз может протекать без участия кислорода (который является конечным акцептором электронов, передаваемых НАДН в дыхательную цепь). При подсчёте энергетического баланса гликолиза следует учитывать, что каждая из реакций второй стадии этого метаболического пути повторяется дважды. Таким образом, в первой стадии было затрачено 2 молекулы АТФ, а во второй стадии путём субстратного фосфорилирования образовалось 2х2 = 4 молекулы АТФ; следовательно при окислении одной молекулы глюкозы в клетке накапливается 2 молекулы АТФ. В нормальном здоровом организме 50% всей молочной кислоты утилизируется печенью, превращаясь в глюкозу. При интенсивной мышечной работе умеренный распад белковых молекул сопровождается выходом аминокислот в кровь и их утилизацией в процессе глюконеогенеза - образованием той же глюкозы. Особенно хорошо утилизируются такиеаминокислоты, как аланин (в печени) и глютаминовая кислота (в кишечнике). От чего же зависит "мощность" глюконеогенеза - основного механизма, "избавляющего" нас от молочной кислоты? От того, насколько интенсивно печень и другие органы синтезируют ферменты глюконеогенеза. 19 (1). Дихотомический аэробный распад глюкозы: схема последо вательности реакций, значение. Энергетический баланс аэробного окисле ния глюкозы. . Аэробным называется окисление биологических субстратов с выделением энергии, протекающее при использовании кислорода в качестве конечного акцептора водорода в дыхательной цепи. В качестве доноров водорода выступают восстановленные формы коферментов (НАДН, ФАДН2 и НАДФН), образующиеся в промежуточных реакциях окисления субстратов. Аэробное дихотомическое окисление глюкозы является основным путём катаболизма глюкозы в организме человека и может происходить во всех органах и тканях. В результате этих реакций глюкоза расщепляется до СО2 и Н2О, а выделяющаяся энергия аккумулируется в АТФ. В этом процессе можно условно выделить три стадии:
9.5.2. На каждом этапе процесса происходит образование восстановленных форм коферментов, которые окисляются ферментными комплексами дыхательной цепи с образованием АТФ путём окислительного фосфорилирования. Коферменты, образующиеся на второй и третьей стадиях аэробного окисления глюкозы, подвергаются непосредственному окислению в митохондриях. В то же время НАДН, образующийся в цитоплазме в реакциях первой стадии аэробного окисления, не способен проникать через митохондриальную мембрану. Перенос водорода с цитоплазматического НАДН в митохондрии происходит при помощи специальных челночных циклов, основным из которых является малат-аспартатный челночный механизм. Цитоплазматический НАДН восстанавливает оксалоацетат в малат, который проникает в митохондрию, где окисляется, восстанавливая митохондриальный НАД; в цитоплазму оксалоацетат возвращается в виде аспартата (рисунок 9.7). 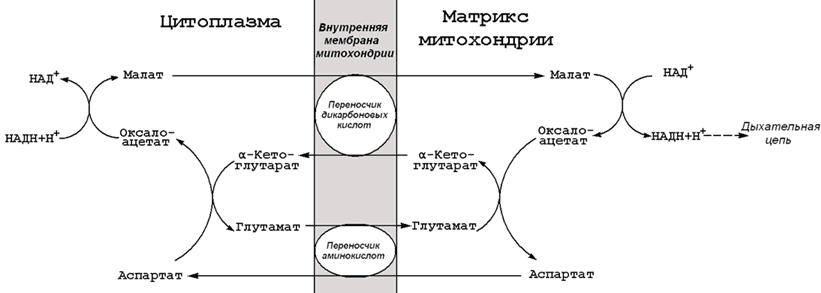 Рисунок 9.7. Малат-аспартатный челночный механизм. Продукция АТФ в реакциях аэробного дихотомического окисления происходит также в трёх реакциях субстратного фосфорилирования – две из них в гликолизе, третья в цикле Кребса на уровне сукцинил-КоА. Полный энергетический баланс аэробного окисления глюкозы представлен на рисунке 9.8. 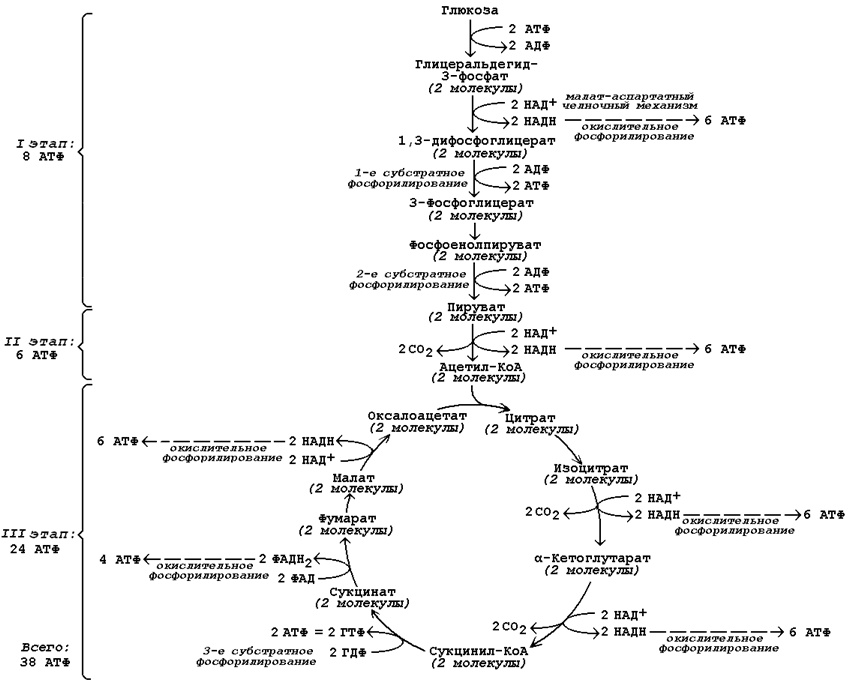 Рисунок 9.8. Энергетический баланс аэробного окисления глюкозы. 20 (1). Пентозофосфатный путь превращений глюкозы. Реакции окисли тельного этапа. Роль пентозофосфатного пути в различных тканях Пентозофосфатный путь представляет собой прямое окисление глюкозы и протекает в цитоплазме клеток. Наибольшая активность ферментов пентозофосфатного пути обнаружена в клетках печени, жировой ткани, коры надпочечников, молочной железы в период лактации, зрелых эритроцитах. Низкий уровень этого процесса выявлен в скелетных и сердечной мышцах, мозге, щитовидной железе, легких. Пентозофосфатный путь называют также апотомическим путём , так как в его реакциях происходит укорочение углеродной цепи гексозы на один атом, который включается в молекулу СО 2 . 15.2.2. Пентозофосфатный путь выполняет в организме две важнейшие метаболические функции :
15.2.3. В пентозофосфатном пути можно выделить две фазы - окислительную и неокислительную. Исходным субстратом окислительной фазы является глюкозо-6-фосфат, который непосредственно подвергается дегидрированию с участием НАДФ-зависимой дегидрогеназы (рисунок 15.3, реакция 1). Продукт реакции гидролизуется (реакция 2), а образующийся 6-фосфоглюконат дегидрируется и декарбоксилируется (реакция 3). Таким образом, происходит укорочение углеродной цепи моносахарида на один углеродный атом ( «апотомия» ), и образуется рибулозо-5-фосфат. 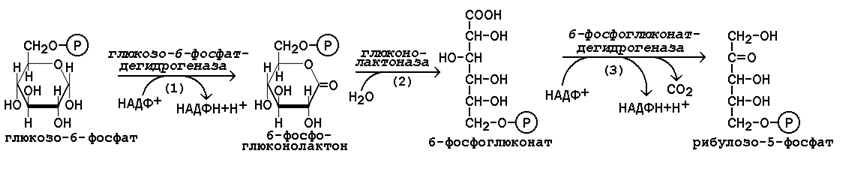 Рисунок 15.3. Реакции окислительной фазы пентозофосфатного пути. 15.2.4. Неокислительная фаза пентозофосфатного пути начинается с реакций изомеризации. В ходе этих реакций одна часть рибулозо-5-фосфата изомеризуется в рибозо-5-фосфат, другая - в ксилулозо-5-фосфат (рисунок 13.4, реакции 4 и 5). 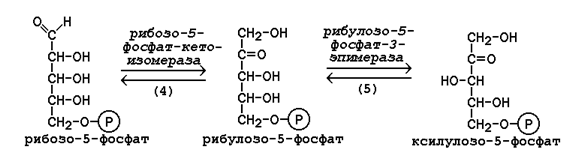 Рисунок 15.4. Реакции изомеризации рибулозо-5-фосфата. Следуюшая реакция протекает при участии фермента транскетолазы, коферментом которой является тиаминдифосфат (производное витамина B 1 ). В этой реакции происходит перенос двухуглеродного фрагмента с ксилулозо-5-фосфата на рибозо-5-фосфат: 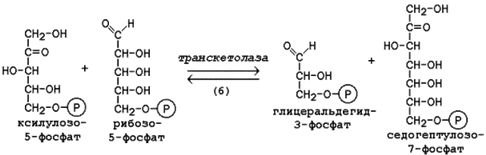 Образовавшиеся продукты взаимодействуют между собой в реакции, которая катализируется трансальдолазой и заключается а переносе остатка дигидроксиацетона на глицеральдегид-3-фосфат. Продукт этой реакции эритрозо-4-фосфат участвует во второй транскетолазной реакции вместе со следующей молекулой ксилулозо-5-фосфата: Таким образом, три молекулы пентозофосфатов в результате реакций неокислительной стадии превращаются в две молекулы фруктозо-6-фосфата и одну молекулу глицеральдегид-3-фосфата. Фруктозо-6-фосфат может изомеризоваться в глюкозо-6-фосфат, а глицеральдегид-3-фосфат может подвергаться окислению в гликолизе или изомеризоваться в дигидроксиацетонфосфат. Последний вместе с другой молекулой глицеральдегид-3-фосфата может образовывать фруктозо-1,6-дифосфат, который также способен переходить в глюкозо-6-фосфат. 15.2.5. Посредством пентозофосфатного пути может происходить полное окисление глюкозо-6-фосфата до шести молекул СО 2 . Все эти молекулы образуются из С-1-атомов шести молекул глюкозо-6-фосфата, а из образовавшихся при этом шести молекул рибулозо-5-фосфата снова регенерируются пять молекул глюкозо-6-фосфата:  Если упростить представленную схему, то получится: Таким образом, полное окисление 1 молекулы глюкозы в пентозофосфатном пути сопровождается восстановлением 12 молекул НАДФ. 21 (1). Обмен гликогена. Регуляция синтеза и распада гликогена. Мобилизация гликогена печени: последовательность реакций, каскадный механизм активации фосфо рилазы. Нарушения обмена гликогена. . Гликоген – биополимер, состоящий из остатков глюкозы, он является компонентом всех тканей животных и человека (см. рисунок 10.1). Этот полисахарид служит основным источником энергии и резервом углеводов в организме. Содержание гликогена в различных органах зависит от физиологического состояния организма. Наиболее высокое содержание гликогена обнаруживается в печени (от 2 до 6% от массы органа). Хотя концентрация гликогена в мышцах значительно ниже (от 0,5 до 1,5%), однако в норме его количество в мышцах составляет 2/3 от общего его содержания в организме. Гликоген отличается значительным разнообразием по структуре и по относительной молекулярной массе. Молекулы его ветвисты. Глюкозные остатки образуют цепи, в которых они связаны между собой α-1,4-гликозидными связями. Разветвления образуются при помощи α-1,6-гликозидных связей. Большая часть глюкозы, поступающей в организм с пищей, превращается в клетках печени в гликоген. Это связано с тем, что накопление легко растворимой глюкозы в клетках привело бы к резкому увеличению осмотического давления и разрушению клеточной мембраны. 12.2.2. Молекула глюкозы фосфорилируется при участии фермента глюкокиназы (в гепатоцитах) или гексокиназы (в других клетках) с образованием глюкозо-6-фосфата. Этот метаболит под влиянием фосфоглюкомутазы превращается в глюкозо-1-фосфат (рисунок 12.4, реакция 1). 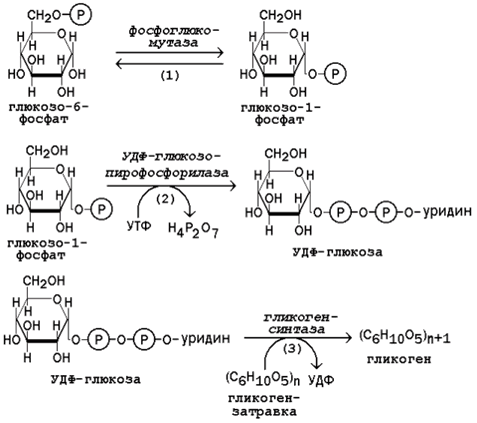 Рисунок 12.4. Реакции биосинтеза гликогена. Глюкозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ) в присутствии фермента УДФ-глюкозо-пирофосфорилазы с образованием УДФ-глюкозы – ключевого метаболита углеводного обмена (рисунок 12.4, реакция 2). Перенос глюкозного остатка с УДФ-глюкозы на затравочную полисахаридную цепь осуществляет гликогенсинтаза – регуляторный фермент данного метаболического пути (рисунок 12.4, реакция 3). Гликогенсинтаза существует в двух формах: гликогенсинтаза b (неактивная) и гликогенсинтаза а (активная). Превращение формы а в форму b катализируется цАМФ-зависимой протеинкиназой, обратный процесс – протеинфосфатазой. Образование α-1,6-гликозидных связей в молекуле гликогена катализирует ветвящий фермент или трансглюкозидаза (рисунок 12.5) . 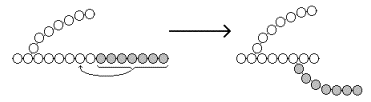 Рисунок 12.5. Образование α-1,6-гликозидных связей в молекуле гликогена Мобилизация, или распад гликогена – процесс превращения гликогена в глюкозу, происходящий в печени. Таким образом, распад гликогена в печени наряду с глюконеогенезом принимает участие в поддержании уровня глюкозы в крови. 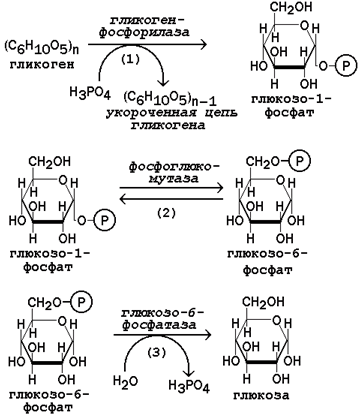 Рисунок 12.6. Реакции мобилизации гликогена. Первую реакцию внутриклеточного расщепления гликогена катализирует фермент фосфорилаза гликогена (рисунок 12.6, реакция 1). Простетической группой его является пиридоксальфосфат (производное витамина В6). Фосфорилаза гликогена – регуляторный фермент, он существует в двух формах: фосфорилаза а (активная) и фосфорилаза b (неактивная). Переход фосфорилазы b в фосфорилазу а катализирует киназа фосфорилазы, которая фосфорилирует неактивный фермент. Превращение фосфорилазы а в фосфорилазу b катализирует протеинфосфатаза, которая осуществляет дефосфорилирование активного фермента. Следует отметить, что ускорение процессов мобилизации гликогена происходит одновременно с торможением его биосинтеза, и наоборот. Активацию фосфорилазы гликогена вызывают гормоны адреналин и глюкагон. Фосфорилаза расщепляет в молекуле гликогена лишь α-1,4-гликозидные связи. Расщепление α-1,6-гликозидных связей осуществляется амило-1,6-гликозидазой гидролитически с образованием свободной глюкозы. Глюкозо-1-фосфат, образующийся в фосфорилазной реакции, при участии уже известного Вам фермента фосфоглюкомутазы переходит в глюкозо-6-фосфат (рисунок 12.6, реакция 2). 12.3.2. Дальнейшая судьба глюкозо-6-фосфата, образовавшегося при распаде гликогена, в разных тканях различна. Как уже упоминалось, в печени и почках имеется фермент глюкозо-6-фосфатаза, отщепляющая фосфат от глюкозо-6-фосфата (рисунок 12.6, реакция 3). Образовавшаяся глюкоза диффундирует в кровь, откуда поглощается клетками организма и служит для них источником энергии. В мышечной ткани фермент глюкозо-6-фосфатаза отсутствует. Поэтому глюкозо-6-фосфат, образовавшийся при распаде гликогена, подвергается дальнейшему превращению по гликолитическому пути с образованием лактата (этот процесс называется гликогенолизом). Образующийся в реакциях субстратного фосфорилирования АТФ используется в процессах мышечного сокращения. Таким образом, гликоген мышц является резервом энергии только для мышечной ткани. 12.3.3. Гликогенозы - наследственные заболевания, характеризующиеся отложением в тканях либо аномально больших количеств гликогена, либо необычных его видов. Такие нарушения содержания и структуры гликогена обусловлены врождённым дефицитом ферментов, участвующих в метаболизме гликогена. Примерами гликогенозов могут служить:
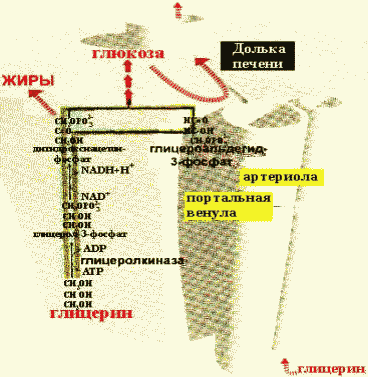
Гликогеноз VI типа (болезнь Херса) – дефицит фосфорилазы в печени. Для этого заболевания характерно повышение содержания гликогена в печени, гипогликемия. После введения адреналина или глюкагона содержание лактата в крови не увеличивается (в отличие от гликогеноза I типа). 22 (1). Биосинтез глюкозы (глюконеогенез): возможные предшест венники, последовательность реакций, значение. Регуляция глюко неогенеза из аминокислот. . Глюконеогенез - биосинтез глюкозы из различных соединений неуглеводной природы. Биологическая роль глюконеогенеза заключается в поддержании постоянного уровня глюкозы в крови, что необходимо для нормального энергообеспечения тканей, для которых характерна непрерывная потребность в углеводах. Особенно это касается центральной нервной системы. Роль глюконеогенеза возрастает при недостаточном поступлении углеводов с пищей. Так, в организме голодающего человека может синтезироваться до 200 г глюкозы в сутки. Глюконеогенез быстрее, чем другие метаболические процессы, реагирует на изменения диеты: введение с пищей большого количества белков и жиров активизирует процессы глюконеогенеза; избыток углеводов, наоборот, тормозит новообразование глюкозы. Интенсивные физические нагрузки сопровождаются быстрым истощением запасов глюкозы в организме. В этом случае глюконеогенез является основным путём пополнения углеводных ресурсов, предупреждая развитие гипогликемии. Глюконеогенез в организме тесно связан также с процессами обезвреживания аммиака и поддержанием кислотно-основного баланса. 12.1.2. Основным местом биосинтеза глюкозы de novo является печень. Глюконеогенез протекает также в корковом слое почек. Принято считать, что вклад почек в глюконеогенез в физиологических условиях составляет около 10% глюкозы, синтезируемой в организме; при патологических состояниях эта доля может значительно возрастать. Незначительная активность ферментов глюконеогенеза обнаружена в слизистой тонкого кишечника. 12.1.3. Последовательность реакций глюконеогенеза представляет собой обращение соответствующих реакций гликолиза. Лишь три реакции гликолиза необратимы вследствие происходящих в ходе их значительных энергетических сдвигов: а) фосфорилирование глюкозы; б) фосфорилирование фруктозо-6-фосфата; в) превращение фосфоенолпирувата в пируват. Обход этих энергетических барьеров обеспечивают ключевые ферменты глюконеогенеза. Обратный переход пирувата в фосфоенолпируват требует участия двух ферментов. Первый из них – пируваткарбоксилаза - катализирует реакцию образования оксалоацетата (рисунок 12.1, реакция 1). Коферментом пируваткарбоксилазы является биотин (витамин Н). Реакция протекает в митохондриях. Роль её заключается также в пополнении фонда оксалоацетата для цикла Кребса. Все последующие реакции глюконеогенеза протекают в цитоплазме. Мембрана митохондрий непроницаема для оксалоацетата, и он переносится в цитоплазму в виде других метаболитов: малата или аспартата. В цитоплазме указанные соединения вновь переходят в оксалоацетат. При участии фосфоенолпируваткарбоксикиназы из оксалоацетата образуется фосфоенолпируват (рисунок 12.1, реакция 2). Фосфоенолпируват в результате обращения ряда реакций гликолиза переходит во фруктозо-1,6-дифосфат. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат катализируется фруктозодифосфатазой (рисунок 12.1, реакция 3). Фруктозо-6-фосфат изомеризуется в глюкозо-6-фосфат. Заключительной реакцией глюконеогенеза является гидролиз глюкозо-6-фосфата при участии фермента глюкозо-6-фосфатазы (рисунок 12.1, реакция 4). 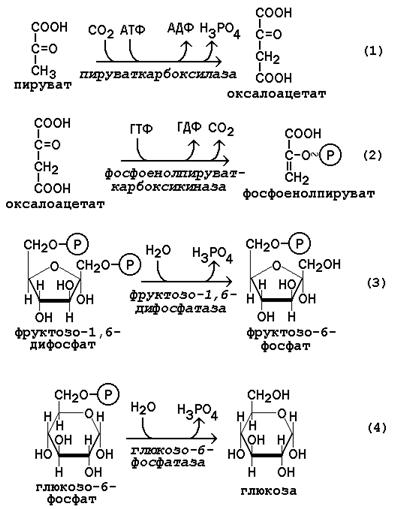 Рисунок 12.1. Обходные реакции глюконеогенеза. 12.1.4. Основными источниками глюкозы в глюконеогенезе являются лактат, аминокислоты, глицерол и метаболиты цикла Кребса. Лактат – конечный продукт анаэробного окисления глюкозы. Может включаться в глюконеогенез после окисления до пирувата в лактатдегидрогеназной реакции (см. раздел «Гликолиз», рисунок 3.4, реакция 11). При продолжительной физической работе основным источником лактата является скелетная мускулатура, в клетках которой преобладают анаэробные процессы. Накопление молочной кислоты в мышцах ограничивает их работоспособность. Это связано с тем, что при повышении концентрации молочной кислоты в ткани снижается уровень рН (молочнокислый ацидоз). Изменение рН приводит к ингибированию ферментов важнейших метаболических путей. В утилизации образующейся молочной кислоты важное место принадлежит глюкозо-лактатному циклу Кори (рисунок 12.2). 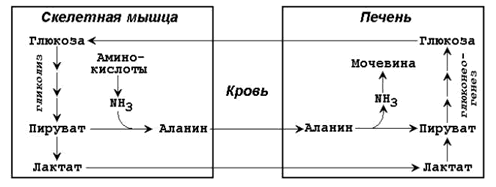 Рисунок 12.2. Цикл Кори и глюкозо-аланиновый цикл (пояснения в тексте). Глюкогенные аминокислоты, к которым относятся большинство белковых аминокислот. Ведущее место в глюконеогенезе среди аминокислот принадлежит аланину, который может превращаться в пируват путём трансаминирования. При голодании, физической работе и других состояниях в организме функционирует глюкозо-аланиновый цикл, подобный циклу Кори для лактата (рисунок 12.2). Существование цикла аланин – глюкоза препятствует отравлению организма, так как в мышцах нет ферментов, утилизирующих аммиак. В результате тренировки мощность этого цикла значительно возрастает. Другие аминокислоты могут, подобно аланину, превращаться в пируват, а также в промежуточные продукты цикла Кребса (α-кетоглутарат, фумарат, сукцинил-КоА). Все эти метаболиты способны преобразовываться в оксалоацетат и включаться в глюконеогенез. Глицерол – продукт гидролиза липидов в жировой ткани.Этот процесс значительно усиливается при голодании. В печени глицерол превращается в диоксиацетонфосфат – промежуточный продукт гликолиза и может быть использован в глюконеогенезе. Жирные кислоты и ацетил-КоА не являются предшественниками глюкозы. Окисление этих соединений обеспечивает энергией процесс синтеза глюкозы. 12.1.5. Энергетический баланс. Путь синтеза глюкозы из пирувата (рисунок 12.3) содержит три реакции, сопровождающиеся потреблением энергии АТФ или ГТФ: а) образование оксалоацетата из пирувата (затрачивается молекула АТФ); б) образование фосфоенолпирувата из оксалоацетата (затрачивается молекула ГТФ); в) обращение первого субстратного фосфорилирования – образование 1,3-дифосфоглицерата из 3-фосфоглицерата (затрачивается молекула АТФ). Каждая из этих реакций повторяется дважды, так как для образования 1 молекулы глюкозы (С6) используются 2 молекулы пирувата (С3). Поэтому энергетический баланс синтеза глюкозы из пирувата составляет – 6 молекул нуклеозидтрифосфатов (4 молекулы АТФ и 2 молекулы ГТФ). При использовании других предшественников энергетический баланс биосинтеза глюкозы отличается. 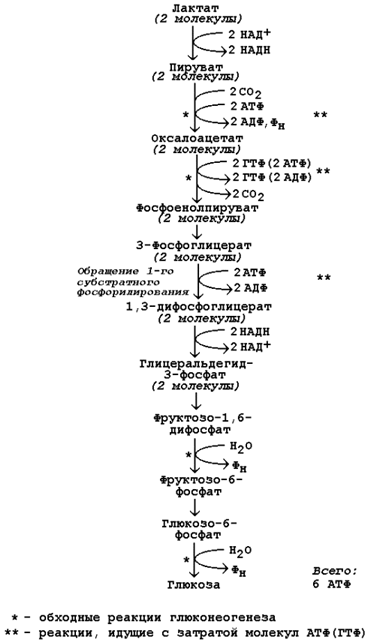 Рисунок 12.3. Энергетический баланс биосинтеза глюкозы из лактата. 12.1.6. Регуляция глюконеогенеза. Скорость глюконеогенеза определяется доступностью субстратов – предшественников глюкозы. Увеличение концентрации в крови любого из предшественников глюкозы приводит к стимуляции глюконеогенеза. Некоторые метаболиты являются аллостерическими эффекторами ферментов глюконеогенеза. Например, ацетил-КоА в повышенных концентрациях аллостерически активирует пируваткарбоксилазу, катализирующую первую реакцию глюконеогенеза. Аденозинмонофосфат, наоборот, оказывает ингибирующее действие на фруктозодифосфатазу, а избыток глюкозы ингибирует глюкозо-6-фосфатазу. Гормон поджелудочной железы глюкагон, гормоны надпочечников адреналин и кортизол повышают скорость биосинтеза глюкозы в организме, увеличивая активность ключевых ферментов глюконеогенеза либо увеличивая концентрацию этих ферментов в клетках. Гормон поджелудочной железы инсулин способствует снижению скорости глюконеогенеза в организме. 23 (1). Роль липидов в организме. Пищевые липиды, суточная потребность. Особенности использования липидов в различных тканях. Депонирование и мобилизация жиров в жировой ткани. Ожирение. Липиды организма человека — это, главным образом, нейтральные сложные эфиры глицерина и высших жирных кислот — триглицериды, фосфолипиды и стерины. Высшие жирные кислоты, входящие в состав сложных липидных молекул в виде углеводородных радикалов, бывают насыщенными и ненасыщенными, содержащими одну и более двойных связей. Липиды играют в организме энергетическую и пластическую роль. По сравнению с молекулами углеводов и белков молекула липидов является более энергоемкой. Поэтому при окислении липидов в организме образуется больше молекул АТФ и тепла. За счет окисления жиров обеспечивается около 50 % потребности в энергии взрослого организма. Запасы нейтральных жиров-триглицеридов в жировых депо человека в среднем составляют 10—20 % массы его тела. Из них около половины локализуется в подкожной жировой клетчатке. Кроме того, значительные запасы нейтрального жира откладываются в большом сальнике, околопочечной клетчатке, в области гениталий и между мышцами. Жиры, откладываясь в жировых депо, служат долгосрочным резервом питания организма. Нарушения обмена липидов у детей приводит к различным расстройствам. Особенно актуально нарушение обмена жиров в жаркое время года, что грозит психическими расстройствами. Жиры являются источником образования эндогенной воды. При окислении 100 г нейтрального жира в организме образуется около 107 г воды. Если в удовлетворении энергетических потребностей организма основную роль играют нейтральные молекулы жира (триглицериды), то пластическая функция липидов в организме осуществляется, главным образом, за счет фосфолипидов, холестерина, жирных кислот. Эти липидные молекулы являются структурными компонентами клеточных мембран (липопротеинов) и предшественниками синтеза стероидных гормонов, желчных кислот и простагландинов. Липиды, г 102г/сут, растительные 72г/сут, незаменимые ж.к, г 3-6, фосфолипиды, г5, растительные липиды, г 20-25 Депонирование и мобилизация жиров Жиры, как и гликоген, являются формами депонирования энергетического материала. Причем жиры — наиболее долговременные и более эффективные источники энергии. При голодании запасы жира у человека истощаются за 5—7 недель, тогда как гликоген полностью расходуется примерно за сутки. Если поступление жира превышает потребности организма в энергии, то жир депонируется в адипоцитах— специализированных клетках жировой ткани. Кроме того, если количество поступающих углеводов больше, чем надо для депонирования в виде гликогена, то часть глюкозы также превращается в жиры. Таким образом, жиры в жировой ткани накапливаются в результате трех процессов:
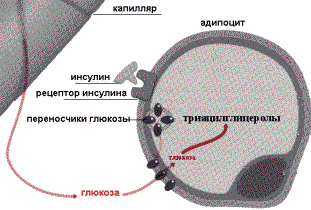
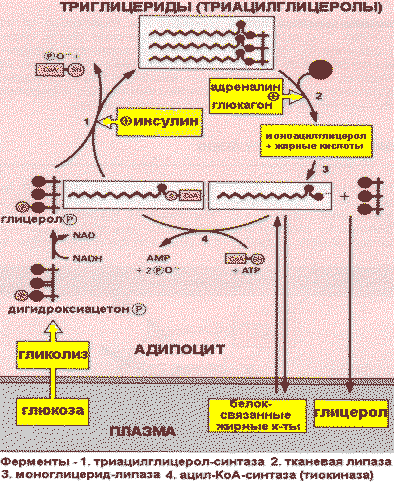
В первом и во втором случае жиры в составе липопротеинов гидролизуются ЛП—липазой и в клетку поступают жирные кислоты, которые затем используются для синтеза ТАГ. Перед включением в ТАГ жирные кислоты сначала активируются путем образования тиоэфиров кофермента А, а затем взаимодействуют с глицеролфосфатом. Так как глицерол не может быть фосфорилирован в адипоцитах (в этих клетках отсутствует глицеролкиназа), то глицеролфосфат образуется при восстановлении диоксиацетонфосфата.
Следовательно, синтез ТАГ может протекать только в присутствии глюкозы, из которой в процессе гликолиза образуется диаксиацетонфосфат. Инсулинстимулирует синтез ТАГ, потому что в его присутствии повышается проницаемость мембран клеток жировой ткани для глюкозы.
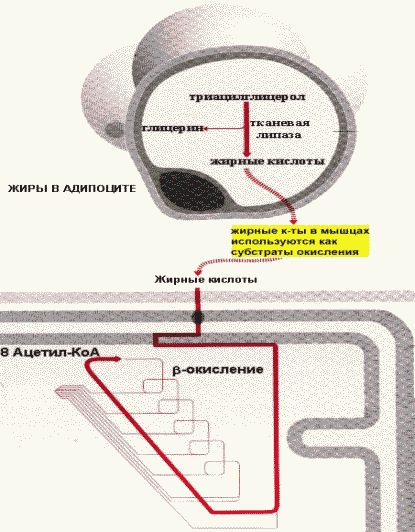
Мобилизацию(липолиз) депонированных ТАГ катализирует тканевая липаза. В результате жиры распадаются на глицерин и свободные жирные кислоты.
Адреналин и глюкагонактивируют внутриклеточную липазу. Действие этих гормонов опосредовано аденилатциклазным каскадом реакций, начиная с активации аденилатциклазы и заканчивая фосфорилированием липазы, которая при этом переходит в активную форму и расщепляет эфирные связи в ТАГ. Глицеролкак растворимое в плазме вещество транспортируется в печень, где используется в реакциях глюконеогенеза. Жирные кислотытранспортируются кровью в виде комплексов с сывороточными альбуминами в разные органы и ткани, где включаются в процесс окисления.
Жировая ткань составляет 20-25% от общей массы тела у женщин и 15-20% у мужчин. Однако избыточное накопление жира в адипоцитах Таблица 8-6. Запасы энергии в организме человека (масса 70 кг)
396 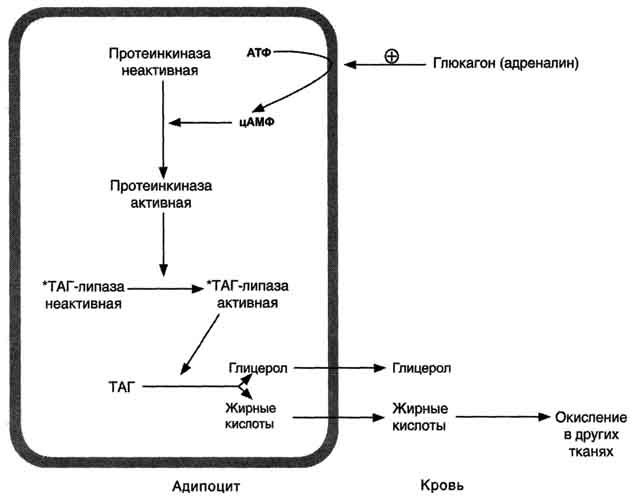 Рис. 8-24. Гормональная регуляция мобилизации жиров в постабсорбтивном периоде, при голодании и физической работе. При голодании увеличивается секреция глюкагона, при физической работе - адреналина. Эти гормоны, действуя через аденилатциклазную систему, стимулируют мобилизацию жиров. *ТАГ-липаза имеет и другие названия: гормончувствительная липаза, тканевая липаза. (ожирение) широко распространено. Среди взрослого населения некоторых стран около 50% людей страдает ожирением. Ожирение - важнейший фактор риска развития инфаркта миокарда, инсульта, сахарного диабета, артериальной гипертензии и желчнокаменной болезни. Ожирением считают состояние, когда масса тела превышает 20% от "идеальной" для данного индивидуума. Образование адипоцитов происходит ещё во внутриутробном состоянии, начиная с последнего триместра беременности, и заканчивается в препубертатный период. После этого жировые клетки могут увеличиваться в размерах при ожирении или уменьшаться при похудании, но их количество не изменяется в течение жизни. Первичное ожирение Первичное ожирение характеризуется множеством гормональных и метаболических особенностей у лиц, страдающих этим заболеванием. В самом общем виде можно сказать, что первичное ожирение развивается в результате алиментарного 397 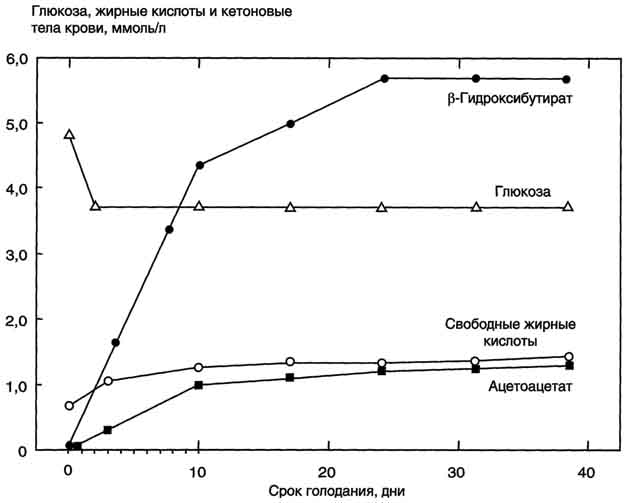 Рис. 8-25. Изменение концентрации жирных кислот, кетоновых тел и глюкозы в крови при голодании. дисбаланса - избыточной калорийности питания по сравнению с расходами энергии. Суточные потребности организма в энергии складываются из:
Затраты энергии, необходимые для физической активности, разделяют на 3 уровня:
В зависимости от интенсивности нагрузки и возраста суточная потребность в энергии колеблется у женщин от 2000 до 3000 ккал в день, а у мужчин - от 2300 до 4000 ккал. Количество потребляемой пищи определяется многими факторами, в том числе и химическими регуляторами чувства голода и насыщения. Эти чувства определяются концентрацией в крови глюкозы и гормонов, которые инициируют 398 чувство насыщения: холецистокинина, нейротензина, бомбезина, лептина. Причины первичного ожирения:
Роль лептина в регуляции массы жировой ткани У человека и животных имеется "ген ожирения" - obese gene (ob). Продуктом экспрессии этого гена служит белок лептин, состоящий из 167 аминокислот, который синтезируется и сек-ретируется адипоцитами и взаимодействует с рецепторами гипоталамуса. В результате его действия снижается секреция нейропептида Y. Нейропептид Y стимулирует пищевое поведение, поиск и потребление пищи у животных. Другие пептиды, участвующие в регуляции чувства сытости, например холецистокинин, также влияют на секрецию нейропептида Y. Таким опосредованным путём лептин выступает регулятором жировой массы, необходимой для роста и репродукции. Уровень лептина у больных ожирением может быть различным. У 80% больных концентрация лептина в крови тучных людей больше в 4 раза, чем у людей с нормальной массой тела. В этих случаях имеется генетический дефект рецепторов лептина в гипоталамусе, поэтому, несмотря на продукцию лептина, центр голода в гипоталамусе продолжает секрецию нейропептида Y. 20% больных имеют изменения в первичной структуре лептина. К настоящему времени описаны 5 одиночных мутаций в гене лептина, которые приводят к развитию ожирения. У этих больных наблюдают повышение отложения жиров в жировой ткани, чрезмерное потребление пищи, низкую физическую активность и развитие сахарного диабета типа II. Патогенез ожирения при дефекте генаob может быть следующим: низкий уровень лептина в крови служит сигналом недостаточного количества запаса жиров в организме; этот сигнал включает механизмы, приводящие к увеличению аппетита и в результате к увеличению массы тела. Следовательно, можно сделать вывод о том, что первичное ожирение - не просто следствие переедания, а результат действия многих факторов, т.е. ожирение - полигенное заболевание. Вторичное ожирение - ожирение, развивающееся в результате какого-либо основного заболевания, чаще всего эндокринного. Например, к развитию ожирения приводят гипотиреоз, синдром Иценко-Кушинга, гипогонадизм и многие другие заболевания 24 (1). Переваривание жиров. Липазы и фосфолипазы. Желчные кислоты и парные желчные кислоты: строение, образование, биологическая роль. Нарушения переваривания липидов. Природные жиры представляют собой смесь триацилглицеринов, различающихся по жирно-кислотному составу. Обычно в жирах обнаруживают смешанные три-ацилглицерины, т. е. содержащие в одной молекуле остатки разных жирных кислот, содержащие в одной молекуле остатки разных жирных кислот, например 1-олеил-2-пальмитил-3-стеарилглицерин, 1,3-диолеил-2-пальмитилглицерин и т. п. В триацилглицеринах человека содержится много ненасыщенных жирных кислот, поэтому жир человека имеет низкую температуру плавления — 10-15 °С; таким образом, в клетках он находится в жидком состоянии. Жиры нерастворимы в воде, и с этим связан ряд особенностей их обмена, в частности необходимость специальных механизмов транспорта с кровью и лимфой, а также возможность депонирования в клетках, подобно гликогену. Биологическая функция жиров тоже подобна функции гликогена: оба эти вещества служат формами запасания энергетического материала.Жиры, наряду с белками и углеводами, относятся к основным пищевым веществам человека. Суточная потребность в них составляет 50-100 г. Жиры обеспечивают до 50 % потребности организма в энергии. Переваривание жиров происходит в тонком кишечнике. В двенадцатиперстную кишку поступает желчь и сок поджелудочной железы, необходимые для переваривания жиров. В соке поджелудочной железы содержится липаза, гидролизую-щая сложноэфирную связь в триацилглицеринах. Под действием липазы жирные кислоты отщепляются от триацилглицерина одна за другой, сначала от а-углерод-ных атомов, потом от р-углеродного атом. Поскольку жиры нерастворимы в водных средах, а липаза нерастворима в жирах, то гидролиз происходит лишь на поверхности раздела этих фаз и, следовательно, скорость переваривания зависит от площади этой поверхности. Фосфолипаза (англ. phospholipase) — фермент, который гидролизует фосфолипиды. В зависимости от положения гидролизуемой связи в фосфолипиде различают 4 основных класса фосфолипаз: A, B, C и D. |