1. Анаэробное дыхание. Аноксигенная фототрофия прокариот
 Скачать 275.4 Kb. Скачать 275.4 Kb.
|

|
Факультативные анаэробы растут как в присутствии,так и в отсутствие О2.Но в зависимости отусловий роста происходят изменения в их метаболизме. Как правило, при наличии молекулярного кислорода такие микроорганизмы переключаются на окисление субстрата с участием О2, т. е. на аэробное дыхание. К аэротолерантным анаэробам принадлежат многие молочнокислые бактерии, способные расти в присутствии молекулярного кислорода, но при этом их метаболизм остается таким же, как и в анаэробных условиях. Облигатные анаэробы не только не нуждаются для роста в наличии молекулярного кислорода,но длямногих видов он токсичен даже в ничтожно малой концентрации. 29. Отношение микроорганизмов к температуре. Различают стенотермные и эвритермные организмы; первые имеют узкий температурный диапазон, вторые — широкий, причем способны приспосабливаться к изменению температуры. Классификация организмов в зависимости от температуры включает следующие категории: Психрофилы —растут в диапазоне от0до20 °С,с оптимумом ниже20 °С.Расти при низкойтемпературе могут также организмы, получившие название психроактивных. Мезофилы —имеют оптимум,близкий к температуре тела теплокровных животных,однако многие лучшерастут при 20–25 °С. Максимальная температура мезофилов около 45 °С. Термофилы —представлены организмами,растущими при температуре выше50 °С.В свою очередь,они делятся на несколько групп. Собственно термофилы имеют максимум развития при температуре ниже 70 °С (62–65 °С) и минимум — выше 40 °С. Экстремальные термофилы имеют максимум при температуре около 90 °С, не растут при 60–70 °С. Наконец, гипертермофилы (90–108 °С) имеют максимум при температуре выше 100 °С. 30. Паракристаллический поверхностный S-слой. S-слой (от англ. surface –поверхность) (паракристаллический поверхностный слой, «поверхностныйS-слой») S-слой представляет собой регулярно построенный поверхностный слой белков или гликопротеинов, расположенный над клеточной стенкой и целиком покрывающий клетку. Последовавшие интенсивные исследования показали, что S-слои встречаются во всех исследованных родах бактерий и архей. Встречается часто, но большинство бакт. и архей его не образуют. Толщина S-слоя - 5-25 нм. Кристаллические решетки S-слоев бактерий и архей могут иметь различные типы симметрии. В зависимости от типа решетки одна морфологическая единица S-слоя содержит одну, две, три, четыре или шесть белковых субъединиц (протомеров). Расстояние между центрами субъединиц - 2.2 - 35 нм Белковые субъединицы соединены в S-слоях нековалентными связями. Под действием агентов, разрушающих водородные связи (при высоких концентрациях солянокислого гуанидина, мочевины и др.), S-слои диссоциируют на субъединицы. S-слой пронизан регулярно расположенными одинаковыми порами. Диаметр пор - 2-8 нм, причем в одном и том же слое могут содержаться поры разных типов. Поры занимают от 30% до 70% площади S-слоя. Две стороны S-слоя неодинаковы: обычно, сторона, обращенная наружу, более гидрофильна, а сторона, прилегающая к клеточной стенке, более гидрофобна. Наружная сторона S-слоя сглажена по сравнению с внутренней, более структурированной стороной. За небольшим исключением, S-слои всех бактерий и архей состоят из какого-либо одного белка (гликопротеина). Белки S-слоя составляют 10-15% общего белка бактериальной клетки, что указывает на чрезвычайно интенсивный синтез и секрецию этих протеинов. На небольшом числе примеров показано, что такой интенсивный синтез связан с наличием сильных промоторов, стабильностью мРНК и использованием предпочтительных кодонов. Функции S-слоев: Широкое распространение S-слоев и высокие затраты клеток на их поддержание (доминантный белок) позволяют думать, что S-слои нужны клеткам (выполняют жизненно важную роль). Но роль S-слоев в целом не ясна.
31. Питание прокариот. Химический состав прокариотной клетки. Макроэлементы и микроэлементы. Пищевые потребности микроорганизмов в соединениях углерода и азота. Факторы роста. Известны два способа питания живых существ:
Разнообразные вещества, в которых нуждаются микроорганизмы и которые потребляются для синтеза основных органических веществ клетки, роста, размножения и для получения энергии называются питательными веществами, а среда,содержащая питательные вещества,называется питательной средой. Потребности микроорганизмов в питательных веществах разнообразны, но независимо от потребностей в питательной среде должны содержаться все необходимые элементы, которые имеются в клетках микроорганизмов, причем соотношение органогенных элементов должно примерно соответствовать этому соотношению в клетке. Источниками водорода и кислорода являются вода, молекулярный водород и кислород, а также химические вещества, содержащие эти элементы. Источниками макроэлементов являются минеральные соли (калий фосфорнокислый, магний сернокислый, железо хлорное и др.). Источниками углерода и азота могут быть как органические, так и неорганические соединения. В соответствии с принятой классификацией микроорганизмов потипу питания их разделяют на группы
В зависимости от источника углерода микроорганизмы делятся на: - автотрофы (сами себя питающие),которые используют углерод из неорганических соединений(углекислого газа и карбонатов); - гетеротрофы (питаются за счет других) –используют углерод из органических соединений.
- фототрофы –микроорганизмы,которые в качестве источника энергии используют энергиюсолнечного света; - хемотрофы – энергетическим материалом для этих микроорганизмов являются разнообразныеорганические и неорганические вещества.
на: - литотрофы – окисляют неорганические вещества и за счет этого получают энергию; - органотрофы –получают энергию путем окисления органических веществ. ганизмов, когда для получения энергии, необходимой для синтеза веществ клетки из диоксида углерода, помимо световой энергии, используется энергия окисления органических соединений. Хемолитоавтотрофия –тип питания,при котором микроорганизмы получают энергию за счет окислениянеорганических соединений, а источником углерода являются неорганические соединения. Хемоорганогетеротрофия – тип питания микроорганизмов,получающих энергию и углерод изорганических соединений. Микроорганизмы, встречающиеся в пищевых продуктах, имеют именно такой тип питания. Среди хемоорганогетеротрофов различают сапрофиты– такие микроорганизмы, которые питаются за счет органических веществ из различных субстратов растительного и животного происхождения (из отмерших клеток и тканей) и паразиты– используют органические вещества из живых клеток. Кроме углерода, важнейшим элементом питательной среды является азот. Автотрофы обычно используют азот из минеральных соединений, а гетеротрофы, кроме неорганических соединений азота, используют аммонийные соли органических кислот, аминокислоты, пептоны и другие соединения. Некоторые гетеротрофы усваивают атмосферный азот (азотфиксаторы). Существуют микроорганизмы, которые сами не способны синтезировать то или иное органическое вещество (например, аминокислоты, витамины). Такие микроорганизмы называют ауксотрофными по данному веществу. Вещества, которые добавляют для ускорения роста и обменных процессов называют ростовыми веществами. Химический состав микробной клетки Основную часть микробной клетки составляет вода (80–90% общей массы клетки). Вода в клетке содержится в свободном и связанном состоянии. Связанная вода входит в состав коллоидов клетки и с трудом высвобождается из них.С потерейсвязанной воды нарушаются клеточные структуры и наступает гибель клетки. Свободная вода участвует в биохимических реакциях, служит растворителем для различных соединений, образующихся в клетке в процессе обмена веществ. При удалении свободной воды гибели клетки не происходит. Важнейшими химическими элементами, преобладающими в клетках микроорганизмов, являются органогенные элементы – кислород,углерод,азот и водород.Из этих элементов состоят органическиевещества (составляют до 85–95% сухих веществ клетки).
Органические вещества клетки представлены: • белковыми веществами. Они состоят из тех же аминокислот, что и белки животных и растений. Наибольшее значение из них имеют нуклеопротеиды – белки, связанные с нуклеиновыми кислотами, которые являются обязательными компонентами ядра и рибосом. Некоторые белки являются ферментами – катализаторами биохимических реакций; • нуклеиновыми кислотами. ДНК содержится в ядре или нуклеоиде, РНК – в ядре, цитоплазме и рибосомах; • углеводами. Они входят в состав различных мембран клетки. Используются для синтеза различных веществ в клетке и в качестве энергетического материала. В клетках микроорганизмов углеводы встречаются преимущественно в виде полисахаридов – гликогена, гранулезы, декстринов, клетчатки. Имеются полисахариды, связанные с белками, липидами; • липидами. Входят в состав цитоплазматической мембраны, а также откладываются в виде запасных питательных веществ. Кроме того, в клетках микроорганизмов содержатся витамины, пигменты и другие органические вещества. Минеральные вещества клетки представлены сульфатами, фосфатами, карбонатами, хлоридами и др. 32. Плазмиды бактерий: форма, размеры, важнейшие свойства, строение. Виды плазмид. Несовместимость плазмид. Плазмида (plasmid):способный к автономной(независимой от основной хромосомы)репликациивнехромосомный генетический элемент, существующий у многих видов бактерий, обычно дающий преимущество клетке-хозяину (например, устойчивость к антибиотикам, тяжелым металлам и т.п.). Это линейные или кольцевые ковалентно замкнутые молекулы ДНК, содержащие от 1500 до 40000 пар нуклеотидов. Для них характерно стабильное существование и наследование в бактериях в ряду клеточных поколений. Используются в качестве векторов для клонирования. Размеры плазмид варьируют от нескольких тысяч до сотен тысяч пар оснований, а число копий на клетку - от одной до нескольких сотен . Хотя многие плазмиды дают клеткам-хозяевам ощутимые селективные преимущества, большинство из них являются криптическими , т.е. не проявляющимися в фенотипе клеток. Рассмотрим классификацию плазмид, основанную на свойствах клетки бактерий:
Плазмидная несовместимость —механизм регуляции числа копий плазмид разного типа вбактериальной клетке. П.н. определяет невозможность внутриклеточного сосуществования близкородственных плазмид. Если плазмиды не могут стабильно сосуществовать в одной клетке в условиях отсутствия селективного давления, их называют несовместимыми. Несовместимость плазмид обусловливается подавлением репликации одной из них и (или) блокированием распределения дочерних молекул ДНК по клеткам перед их делением. Эти оба механизма действуют независимо друг от друга. 33. Поверхностные органеллы: целлюлосомы, шипы и экстрацеллюлярные газовые баллоны. Шипы (или «спин» от лат. spina-шип) Распространенность:Обнаружены у грам(-) (в частности,у псевдомонад,метилотрофов,цианобактерий)и не обнаружены среди архей. Строение: Жесткие полые белковые цилиндры,отходящие перпендикулярно от кл.стенки(прикрепленык наружной поверхности внешней мембраны). Шипы не протыкают насквозь оболочку клеток. У основания цилиндры могут расширяться раструбом. В среднем на 1 кл. – 10 шипов, расположены нерегулярно (беспорядочно, случайно). Длина шипов – 0,5-3 мкм, диаметр – 50-100 нм. Цилиндр образован одним белком - спинином, субъединицы которого уложены в спираль. Функции в целом неизвестны(хотя на обр-е шипов расходуется много белка кл.).Бакт.с шипами обычнонеподвижны. Считают, что обр-е шипов способствует лучшему выживанию бакт. в естеств. среде обитания. Предположительно они способствуют: 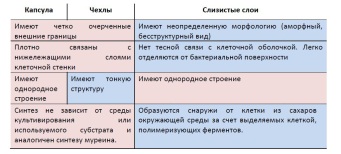
|