1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
1. О сущности живого. Нуклеопротеидные комплексы.Эволюция представлений о химической сущности жизни. Ф.Энгельс: «Жизнь – способ существования белковых тел» Жизнь – активная форма существования материи; период существования отдельно взятого организма от момента его возникновения до старости. Нач XX в. академик Кольцов – гипотеза «Особых кольцевых молекул белков» ДНК как хим соед-е идентифицировано ещё в XIX в. Мишер. Опыт Гриффитса 1926 – феномен трансформации (в феномене трансформации два участника: бакт и чужеродн ДНК, к-ая измен св-ва бактерии. ТФ – трансформирующий фактор - из убитого S-штамма вызвал превр-е R-штамма в S-штамм) Гриффитс не смог определить химическую природу ТФ. 1944-лаб-я Эвери – экспериментальные доказательства - ТФ идентичен ДНК. 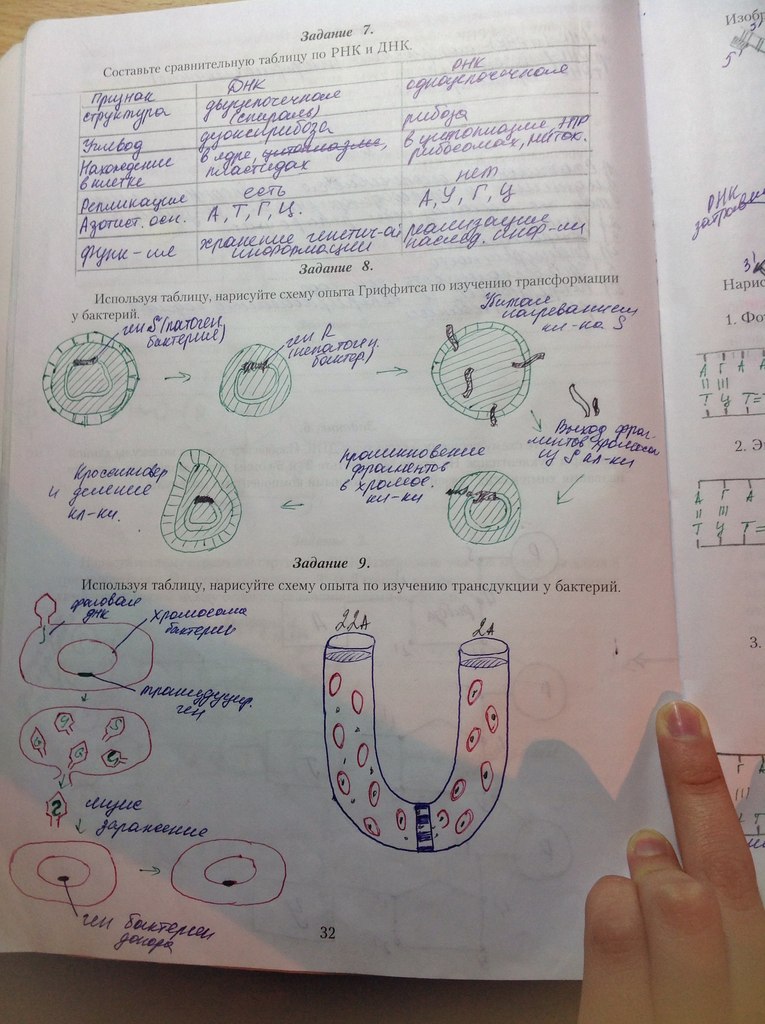 R + мышь – жив; S + мышь – мёртв; S(t) + мышь – жив; S(t) + R - мёртв В живых системах 3 потока: ЭНЕРГИИ, ВЕЩЕСТВА и ИНФОРМАЦИИ, кот. подчиняются законам термодинамики. 1 ЗАКОН: В плане энергии нельзя выиграть (переходит из 1 вещ в другое) 2 ЗАКОН: В плане энергии нельзя остаться «при своих» (при переходе энергии ее часть теряется, выделяется в виде тепла) Нуклеи к-ты (ДНК, РНК) и белки являются субстратом жизни. Ни нуклеин к-ты, ни белки в отдельности не являются субстратами жизни. Поэтому считают, что субстратами жизни являются нуклеопротеиды. Нет живых систем, не содержащих их (от вирусов до человека). Однако они являются субстратом жизни лишь когда находятся и функционируют в клетке, Вне клеток – это обычные химич соед-я. Следоват-но, жизнь – это взаимод-е нуклеин к-т и белков, а живое – то, что содержитсамовоспроизводящуюся молекулярную систему в виде механизма активного воспроизв-ва синтеза нуклеиновых кислот и белков. Жизнь существует в виде нуклеопротеидных комплексов. 2. Клетка – миниатюрная биосистема. 5 признаков живых систем. (см 1 вопр) Клетка – это самостоятельная биосистема, уровень организации живой материи, кот присущи проявления основных свойств живого: 5 признаков живых систем: 1. Открытость (живые системы обмениваются с окр средой энергией, веществами, информацией) 2. Самообновление (системы эволюционируют во времени) 3. Саморегуляция (гомеостаз; системы не требуют регуляции из вне) 4. Самовоспроизведение 5. Высокоупорядоченность Клетка представляет собой единицу строения, развития и размножения организмов -самоуправляемая система. Управляющая генетическая система клетки предствалена сложными макромолекулами - нуклеиновыми кислотами (ДНК и РНК). Клетка может существовать только как целостная система, неделимая на части. Целостность клетки обеспечивают биологические мембраны. Клетка - элемент системы более высокого ранга - организма. Части и органоиды клетки, состоящие из сложных молекул, представляют собой целостные системы более низкого ранга. Клетка рассматривается в качестве общего структурного элемента живых организмов. Клеточня теория – одно из общепризнанных биологических обобщений, утверждающих единство принципа строения живых организмов. Современная клеточная теория включает следующие основные положения: 1. Клетка – единица строения (все живые существа состоят из клеток). 2. Клетка – единица жизнедеятельности (все клетки сходны по строению, химическому составу и жизненным функциям). 3.Клетка – мельчайшая единица живого (каждая клетка реализует все св-ва живого) 4. Клетка – единица размножения (кажд клетка возникает из клетки) – Р.Вирхов 3. Клетка – элементарная еденица живого. Отличительные признаки про- и эукариотических клеток. Клетка – элементарная единица живого, основная единица строения, функционироваия, размножения и развития всех живых организмов. Клетка представляет собой биосистему, которой присущи все признаки живых систем.
4. Принцип компартментации. Биологическая мембрана. 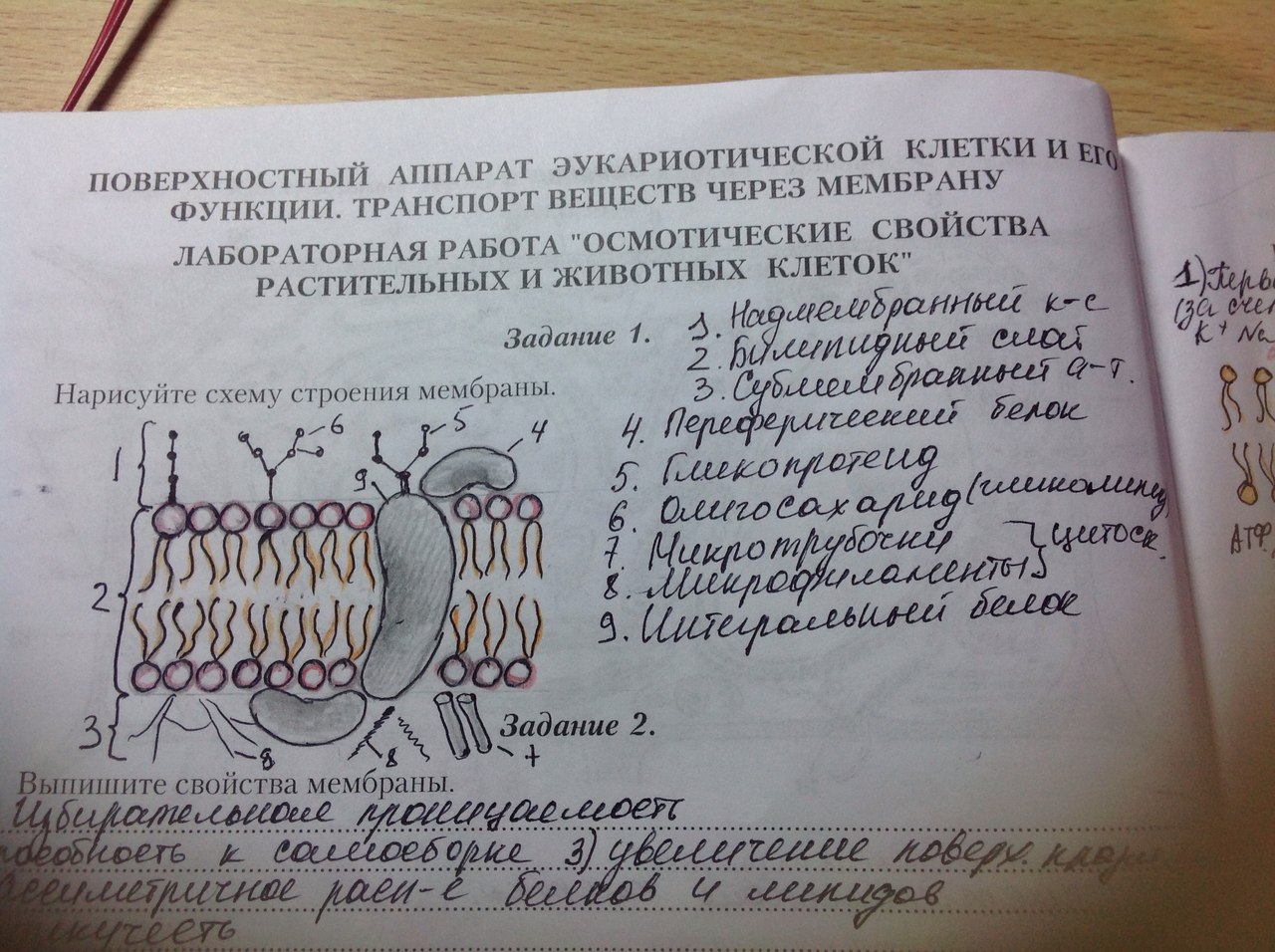 Высокая упорядоченность внутреннего содержимого клетки достигается путём компартментации её объёма – подразделения на отсеки, отличающиеся деталями хим.состава. Компартментация – пространственное разделение веществ и процессов в клетке. Компартменты – отсеки, ячейки – ядро, митохондрия, пластиды, лизосомы, вакуоли, т.к. образ мембраны. Рис. 2.3. Компартментация объема клетки с помощью мембран: 1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4—транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 — гранула секрета 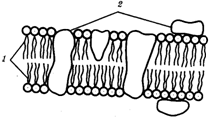 Билипидный слой – гидрофобные хвосты – внутрь, гидрофильные головки – наружу. Мембранные белки:
Мембранные липиды:
Ф-ции мембраны: барьерная (защищает внутр содерж-е клетки), поддерживает постоянную форму кл-ки; обеспечивает связь клеток; пропускает внутрь кл-ки необходимые в-ва (избират прониц-ть – мол-лы и ионы проходят через мембрану с различной скоростью, чем больше размер, тем меньше скор-ть). Свойства мембраны: - билипидный слой способен к самосборке; - увелич-е пов-ти мембраны за счёт встраивания в неёмембранных пузырьков (везикул); - белки и липиды ассиметрично расположены в плоскости мембраны; - белки и липиды могут перемещаться в плоскости мембраны в пределах слоя (латеральное перемещ-е); - наружн и внутр пов-ти мембраны имеют разный заряд. - мембрана обеспечивает разделение заряженных частиц и поддержание разности потенциалов 5. Принцип клеточной компартментации. Организация и свойства биологической мембраны. История изучения. См. 4 вопрос. История изучения: 1902, Овертон находит липиды в составе пзазматической мебраны. 1925, Гортер и Грендел показывают наличие двойного слоя липидов в мембране эритроцитов. 1935, «бутербродная» модель Даниелли и Давсона (липидный бислой между двумя слоями белков) Накопл-е фактов, необъяснимых с позиции «бутербродной» мембраны (мембраны очень динамичны) 1962, Мюллер создает плоскую модель искусственной мембраны 1957-1963, Робертсон формулирует понятие элементарная биологическая мембрана. 1972, создание Зингером и Николсоном жидкостно-мозаичной модели мембраны. 6. Структурная организация и свойства биологических мембран. См. 5 вопрос 7. Мембранные белки и липиды. Мембранные белки:
Мембранные липиды:
8. Явление осмоса в растительных и животных клетках. 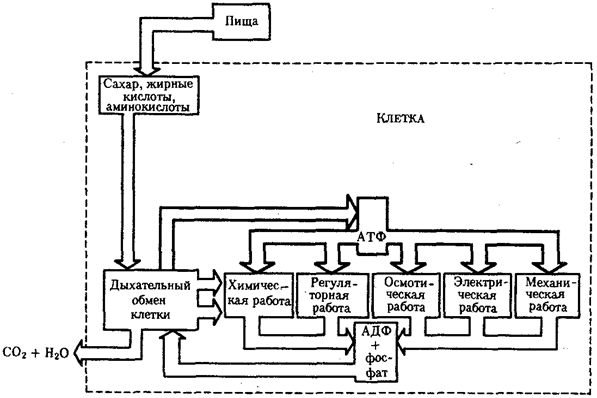 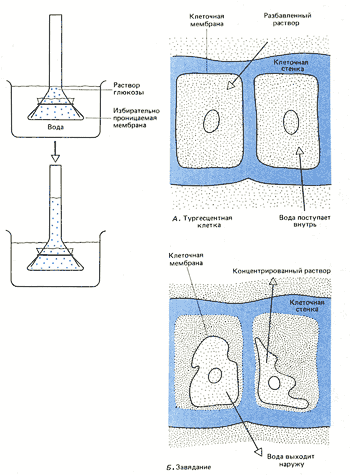 Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы. Одна из них осмотическая (поддержание перепадов концентрации веществ) Осмос - диффузия (передвижение мол-л по градиенту конц-ции - из обл выс конц в обл низк конц) воды через полупрониц мембраны. В раст кл-ке: Плазмолиз (когда жарко) - отток воды, содерж-е кл-ки сжим-ся и отходит от клет стенки. Деплазмолиз (прохл-но+полить) - кл-ки набух и прижим к клет стенке, подчин тургорному давл-ю (тургор - внутр гидростатич давл, вызывающее натяж-е кл стенки). Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором). В жив кл-ке: изотонич р-р - норма, гипертон р-р - сморщив-е, гипотонич р-р - набух-е, затем лопаются-лизис. Рис. 1. Осмос в искусственной системе. Трубку, содержащую раствор глюкозы и закрытую с одного конца мембраной,пропускающей воду, но не пропускающей глюкозу, опускают закрытым концом в сосуд с водой. Вода может проходить через мембрану в том и в другом направлении; однако молекулы глюкозы в трубке мешают движению соседних молекул воды, и потому больше воды входит в трубку, чем выходит из нее. Раствор поднимается в трубке до тех пор, пока давление его столба не станет достаточным для того, чтобы вытеснять воду из трубки с такой же скоростью, с какой она поступает внутрь. 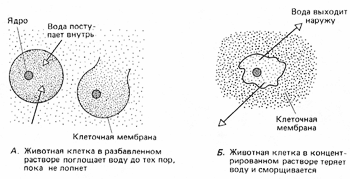 Осмос - процесс одностороннего проникновения молекул растворителя через полунепроницаемую мембрану в сторону большей конц. растворенного вещ-ва. отчего зависит осмос? во-первых, от общей концентрации всех растворенных частиц по обе стороны от мембраны, ну а во-вторых от давления, создаваемого каждым "р-ром" (понятие осмотического давления: такое давление на раствор, обусловленное стремлением системы (ну т.е. клетки) выровнить конц. р-ра в обеих средазх, разделенных мембраной). Наличие воды необходимо для норм. протекания всех ппроцессов, и именно благодаря осмосу происходит "обводнение" клеток и структур. У КЛЕТОК НЕТ СПЕЦ, МЕХАНИЗМА ДЛЯ НАСАСЫВАНИЯ И ОТКАЧИВАНИЯ ВОДЫ НЕПОСРЕДСТВЕННО! - поэтому приток и отток воды регулируется изменением конц. вещ-в. Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором). 9. Особенности строения растительных клеток. Осмотические свойства растительных клеток. Особ строен растит кл-к: жёстк целлюлозопектинов стенка,пластиды, вакуоли с клет соком. Жёсткость кл стенки предотвращ от чрезмерн набух-я и разрыва, обуславл потерю спос-ти к передвиж-ю. За счёт роста вакуоли увел размер кл-ки, игр важн роль в регуляции поступл-я воды в кл-ку,содерж растит антибиотики,к-ые убив микроорг-мы и микроскопич грибы. Пластиды-неоднородн группа органелл растит кл-ки (хлоропл-ты, хромопл-ты и лейкопласты) Фотос-з –синт-з сложн орг в-в из неорг приучаст солн света Свет фаза 1-поглощ-е света хлорофиллом,возбужд-е его е.2-возбужд е перемещ по цепи переноса,отдавая лишн энерг на синтез АТФ 3-фотолиз воды (итог-синтез АТФ+фотолиз воды с выдел О2) Темн фаза 1-улавлив-ся СО2 2-синтез глюкозы из СО2 с пом энерг АТФ Отличие растительной от животной клетки: ВАКУОЛЬ. Окруж.мембраной-стонопласт.Компартмент, связанный с неподвижн обр жизни растит кл+ПЛАСТИДЫ(хлоропласты, хромопласты, лейкопласты) Функции: Накопительная(вода, глю, к-ты, фруктоза)+ненужные и невыводимые вещ-ва Алкалоиды-биологич. Активные вещ-ва; Пигменты(окраска зависит от рН) Поддержание осмотического давления(Тургор) Защитная(Бактериологические св-ва-фитонциды) Ферментативная (роль мезосомы) Нет клеточн.центра! Не способна к фагоцитозу(мешает клет.стенка)! Механич прочность клет.стенок позволяет сущ в гипотонической среде, где в кл ОСМОТИЧЕСКИМ путем поступает вода. По мере поступления воды в клетку, возникает давление, препятствующее дальнейшему пост воды. Избыточное гидростатическое давление в клетке-ТУРГОР-обеспечение роста, сохр формы растением, определ положение в пространстве, противостояние механич воздействиям. 10. Поверхностный аппарат клетки. Пассивный транспорт. 1. Плазматическая мембрана Билипидный слой – гидрофобные хвосты – внутрь, гидрофилльные головки – наружу. Мембранные белки:
Мембранные липиды:
Ф-ции мембраны: барьерная (защищает внутр содерж-е клетки), поддерживает постоянную форму кл-ки; обеспечивает связь клеток; пропускает внутрь кл-ки необходимые в-ва (избират прониц-ть – мол-лы и ионы проходят через мембрану с различной скоростью, чем больше размер, тем меньше скор-ть). 2. Надмембранный комплекс – гликокаликс
Ф-ции: изолирующая, рецепторная, маркировочная, ферментативная 3. Субмембранный комплекс – примембранная часть цитоскелета с белками,обеспечив-щими её связь с мембр) Ф-ции: поддерж-е формы, изменение пов-ти (при фагоцитозе и пиноцитозе) 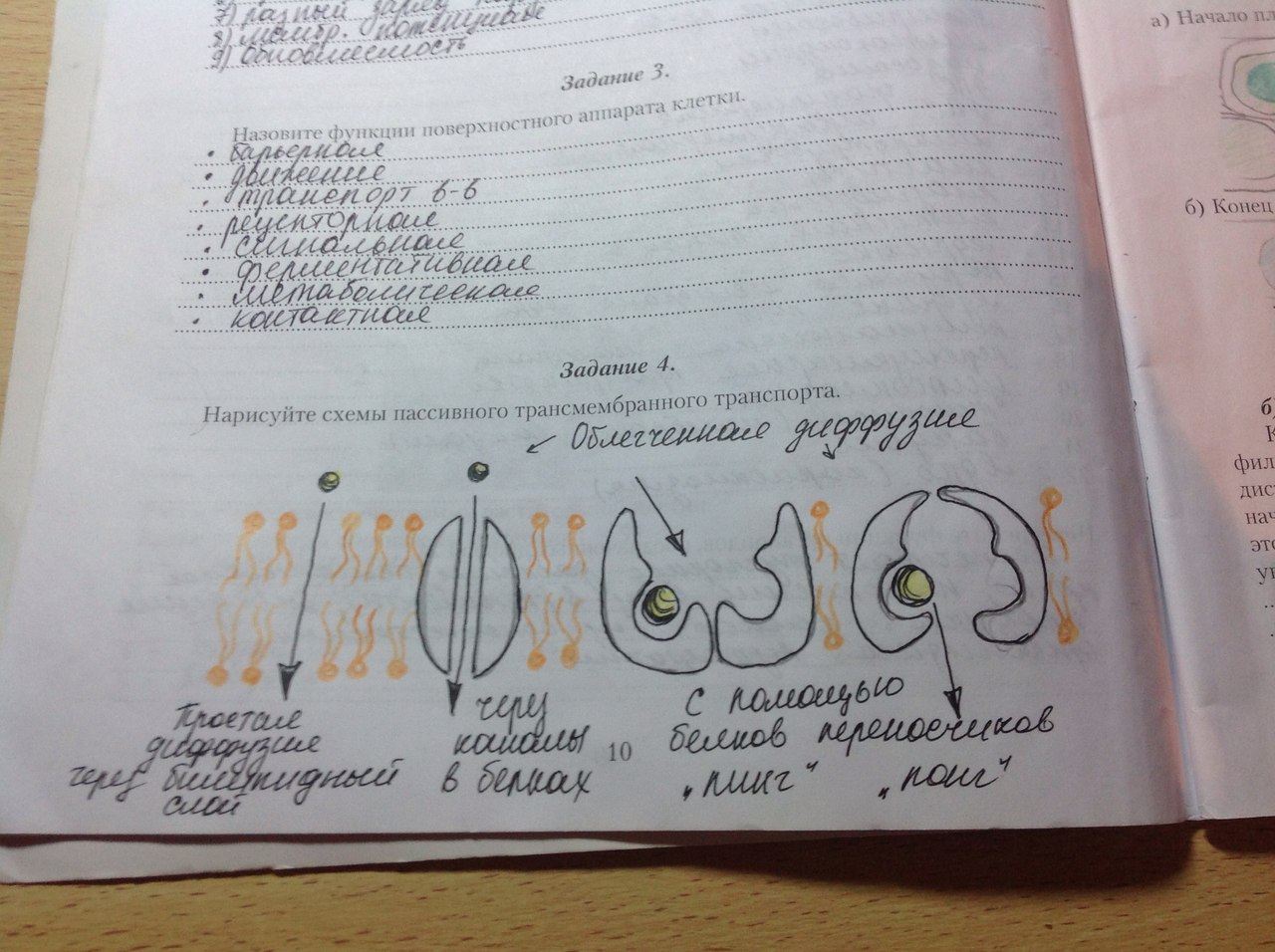 Пассивный транспорт-транспорт вещест, который осуществляется по градиенту концентрации и не требует затрат энергии. Если молекула заряжена, то на ее транспорт влияют как градиент концентрации, так и разница электрических потенциалов со стороны мембраны (мембранный потенциал). Вместе градиент концентрации и электрический градиент составляют электрохимический градиент. Активный и пассивный транспорт осуществляется с участием мембранных белков: белки переносчики (изменяют свою конформацию) и каналообразующие (формируют заполненные водой поры, пронизывающие липидный бислой; такие белковые каналы обладают ионной избирательностью, могут открываться при действии различных факторов, таких как: изменение мембранного потенциала, механическая стимуляция, при связывании с определенным лигандом. Примеры пассивного транспорта: Диффузия-передвижение молекул по градиенту концентрации, которое может происходить либо через билипидный слой (так транспортируются растворимые в липидах неполярные и слобополярные молекулы-кислород, углекислый газ, стероидные гормоны…), либо через поры, образуемые мембранными белками (так, например, перемещаются молекулы воды). Облегченная диффузия-пассивный транспорт, в котором участвует транспортный белок (переносчик или каналообразующий). Белок-переносчик может пребывать в двух конформационных состояниях. В состоянии «пинг» этот белок соединяется с транспортируемой молекулой или ионом. Это вызывает его преход в состояние «понг», в котором он отдает транспортируемое вещество на противоположной стороне мембраны. Освободившись от транспортируемого вещества белок принимает исходное состояние (конформацию). |