1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
54. Поверхностный аппарат ядра. Поровые комплексы. Взаимосвязь ядра и цитоплазмы. 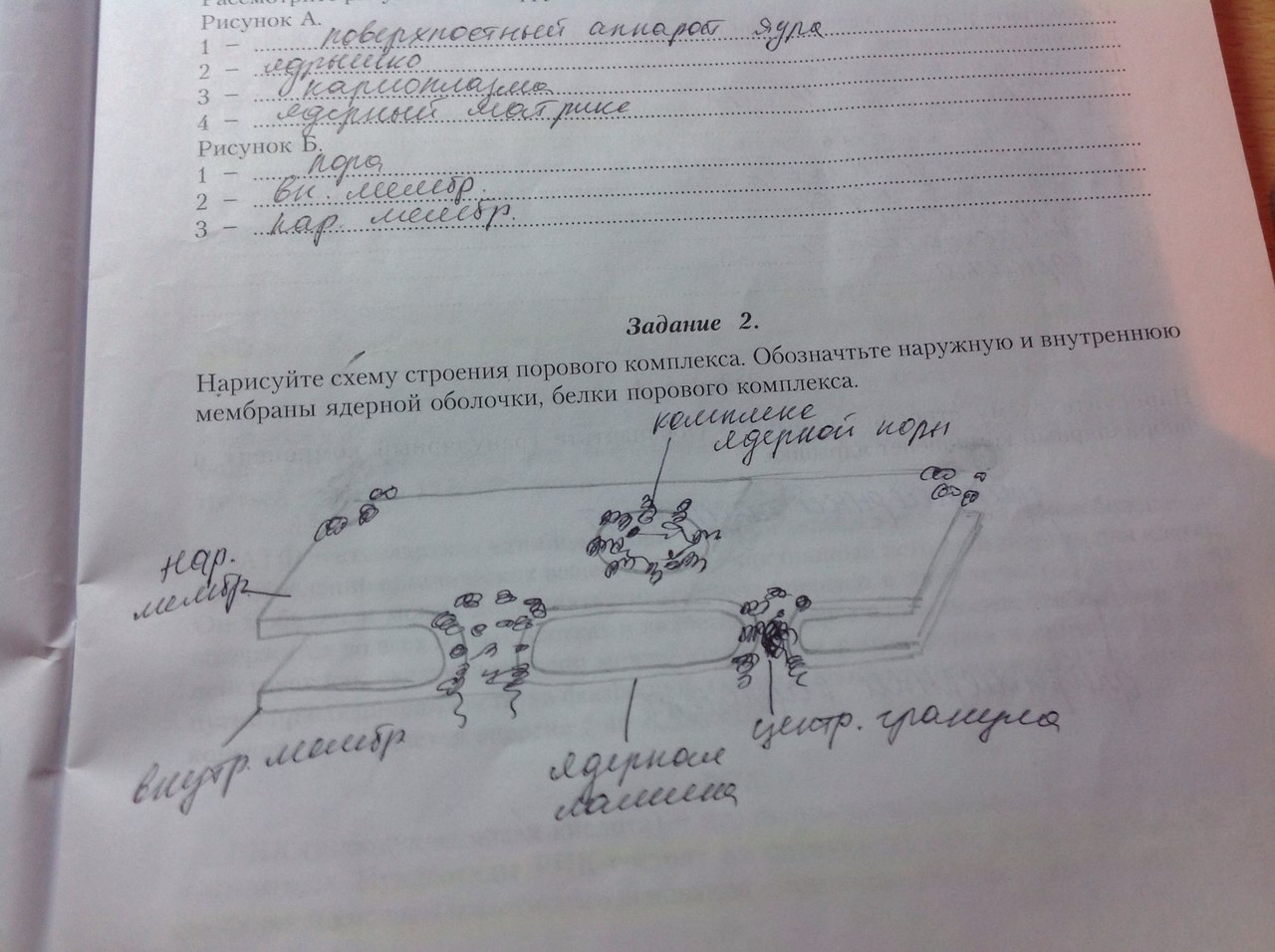 Поверхностный аппарат: ядерная оболочка с поровым комплексом и ламина. Ядерная оболочка образована наружной и внутренней ядерными мембранами, между которыми перинуклеарное пространство. Наружная переходит во внутреннюю в области ядерных пор. Наружная мембрана переходит в мембраны ЭПР, а перинуклеарное пространство таким образом оказывается связанным с полостью каналов и цистерн ЭПР. Поровый комплекс представляет собой 2 кольца из 8 белковых глобул, расположенных по краю порового отверстия- в области слияния наружной и внутренней мембран. В центре поры может быть видна центральная гранула. Поровый комплекс представляет собой надмолекулярную структуру. Они обладают собственными рецепторами, аппаратом, регулирующим направление, способ и интенсивность транспорта через пору. Ламина - плотная пластинка, примыкающая к внутренней мембране ядерной оболочки. Представляет собой густую сеть белковых фибрилл. Ламина способствует поддержанию формы ядра, выполняет функцию упорядочения расположения интерфазных хромосом, связана с цитоскелетом и играет важную роль в поддержании порового комплекса. 55. Структура ядрышка. Ядрышко – источник рибосом. Строение рибосом. Амплификация ядрышек. Ядрышко- представляет собой структуру в которой происходит образование рибосомальных единиц. Здесь находятся участки ДНК, содержащие многочисленные одинаковые гены рРНК. В метафазных хромосомах эти участви(ядрышковые организаторы) локализованы в области вторичной перетяжки. У человека они находятся в 13,14,15,21,22 хромосомах. Кроме того, гены рРНК находятся также в 1 паре хромосом. Различают фибриллярный и гранулярный компоненты ядрышка. Фибриллярная зона содержит ДНК, рРНК, а гранулярная часть-зона со зрелыми субъединицами рибосом. В клетках эукариот существует 2 разновидности рибосом: рибосомы цитоплазмы (80S) и рибосомы, находящиеся в митохондриях и пластидах (50-80S) Рибосомы состоят из двух субъединиц большой и малой. Малая субъединица рибосомы удерживает мРНК и тРНК, а большая катализирует образование пептидной связи. В состав субъединиц входят рРНК и белки (преимущественно глобулярные). Большая субъединица рибосомы состоит из 3 молекул рРНК и 50 белков, а малая из 1 молекулы рРнк и 30 белков. Субъединицы рибосом собираются в ядрышке и через ядерные поры выходят в цитоплазму, где находятся в диссоциированном состоянии. 2 субъединицы образуют комплекс- рибосому только при осуществлении синтеза полипептида на иРНК(процесс трансляции). Если к иРНК прикрепляется множество рибосом, то такой комплекс называется полисомой. Амплификация . Обычно число генов рРНК постоянно на геном, оно не меняется в зависимости от уровня транскрипции этих генов. Так у клеток с высоким уровнем метаболизма число генов рРНК точно такое же как и число у клеток, полностью прекративших синтез рибосом. При репликации ДНК в S-периоде происходит и удвоение числа генов рРНК, поэтому их количество коррелирует с плоидностью клетки. Однако существуют случаи, когда гены рРНК подвергаются избыточной репликации. При этом дополнительная репликация генов рРНК происходит в целях обеспечения продукции большого количества рибосом. В результате такого сверхсинтеза генов рРНК их копии могут становиться свободными, экстрахромосомными. Эти внехромосомные копии генов рРНК могут функционировать независимо, в результате чего возникает масса свободных дополнительных ядрышек, но уже не связанных структурно с ядрышкообразующими хромосомами. Это явление получило название амплификации генов рРНК. 56. Ядро – система хранения, воспроизведения и реализации генетический информации.  Клеточное ядро отделено от содержимого клетки оболочкой. Функции ядра состоят в хранении наследственного материала (ДНК), его воспроизводстве (репликация ДНК) с целью передачи в ряду клеточных поколений (митоз), а также в реализации наследственной информации в ходе биосинтеза белка в жизнедеятельности клетки (транскрипция, процессинг пре-РНК транскриптов). В нем образуются структурные элементы - большая и малая субъединицы - цитоплазматических органелл рибосом, на которых в цитоплазме происходит образование полипептидов (простых белков). В ядре выделяют ядерную оболочку, ядерный матрикс, ядрышко, хромосомы (хроматин), ядерный сок. В ядре сосредоточена большая часть ДНК эукариотической клетки - 90%. Она распределена между ядерными структурами – хромосомами. Морфология хромосом меняется по стадиям клеточного цикла. При вхождении клетки в митоз материал хромосом приобретает плотную упаковку (митотическая форма),а вне митоза - рыхлую (интерфазная форма). хроматин- вещество хромосомы, а его состав и плотность упаковки различаются по длине хромосомы в зависимости от стадии это го цикла. 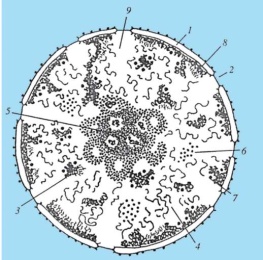 Хромосомы во взаимодействии с внехромосомными механизмами обеспечивают: • хранение генетической информации; • использование этой информации для воспроизводства и поддержания клеточной организации и функций; • регуляцию считывания (транскрипция) наследственной информации; • удвоение (репликация, самокопирование) генетического материала материнских клеток перед клеточным делением; • передачу этого материала дочерним клеткам в процессе митоза. Первую из этих функций хромосома выполняет в обеих структурных формах - митотической и интерфазной, следующие три функции - в интерфазной форме, последнюю - в митотической форме. Хромосомная организация наследственного материала эукариот создает условия для тонкой регуляции генетических функций, репаративных процессов, минимизирующих объем нарушений молекулярной структуры ДНК, а также для рекомбинации ДНК в ходе мейоза при образовании половых клеток (см. кроссинговер, комбинативная геноти-пическая изменчивость). Рис. 2.8.Структура клеточного ядра (схема): 1 - ядерная оболочка (две мембраны - внешняя и внутренняя, и перинуклеарное пространство); 2 - ядерная пора; 3 - конденсированный хроматин; 4 - диффузный хроматин; 5 - ядрышко (гранулярный и фибриллярный компоненты, в центральных светрых зонах находится р-ДНК); 6 - интерхроматиновые гранулы (РНП); 7 - перихрома-тиновые гранулы (РНП); 8 - перихроматиновые фибриллы (РНП); 9 - кариоплазма, ядерный сок 57. Организация эу- и гетерохроматина. Структура и химия хроматина. В ядре сосредоточена большая часть ДНК эукариотической клетки - 90%.в составе хромосом. Материал хромосом - совокупность глыбок, зерен и волоконец – хроматина. Химический состав хроматина (хромосом) эукариотической клетки Большая часть объема хромосом представлена ДНК и белками. Заметные химические компоненты хромосом - РНК и липиды. Среди белков (65% массы хромосом) выделяют гистоновые (60-80%) и не-гистоновые. Также присутствуют полисахариды, ионы металлов (Ca, Mg)и др. Особое место среди хромосомных белков принадлежит гистонам. В составе нуклеогистонового комплексаДНК менее доступна ферментам нуклеазам, вызывающим ее гидролиз (функция защиты). Гистоны выполняют структурную функцию, участвуя в процессе компактизации хроматина. Гистоновые белки представлены пятью видами (фракциями): Н1, Н2А, Н2В, Н3и Н4. Число ядерных негистоновых белковпревышает несколько сотен. Они удерживают «открытую» конфигурацию хроматина, «разрешающую» доступ к биоинформации ДНК, то есть ее транскрипцию. К категории «временных» относятся цитозольные белки-рецепторы (функционально-транскрипционные факторы), захватывающие сигнальные молекулы, в комплексе с которыми они проникают в ядро и их активируют. РНК хромосомпредставлена продуктами транскрипции, еще не покинувшими место синтеза, - непосредственный продукт транскрипции генов или пре-и(м)РНК, пре-рРНК, пре-тРНК транскрипты. Некоторые виды РНК «временного внутриядерного пребывания» создают условия для основного процесса, выполняя сигнальную функцию. Так, репликация ДНК требует для своего начала образуемой «на месте» РНК-затравки (РНК-праймер), которая по завершении процесса разрушается здесь же в ядре. В зависимости от степени компактизации материал интерфазных хромосом представлен эухроматином и гетерохроматином . Эухроматин - низкая степень компактизации и неплотная «упаковка» хромосомного материала. Эухроматин представлен, в основном, ДНК с уникальными последовательностями нуклеотидов. Гены из эухроматизированного участка хромосомы, оказавшись в гетерохроматизированномучастке или рядом с ним, обычно инактивируются. Гетерохроматинотличается высокой степенью компактизации, то есть плотной «упаковкой» материала хромосомы. Большая его часть представлена умеренно или многократно повторяющимися нуклеотидными последовательностями ДНК. К первым относятся мультикопийные гены гистонов, рибосомных и транспортных РНК. 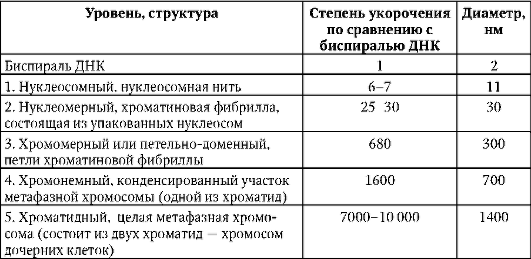 58. Уровни структурной организации хроматина. Компактизация хроматина. На протяжении клеточного цикла хромосома сохраняет структурную целостность благодаря компактизации-декомпактизации (конденсация-деконденсация)хромосомного материала – хроматина. Вследствие компактизации при переходе хромосом из интерфазной формы в митотическую суммарный линейный показатель сокращается примерно в 7-10 тыс. раз. Таблица 2.1.Последовательные уровни компактизации хроматина. В образовании нуклеосомной нити ведущая роль принадлежит гистонам Н2А, Н2В, Н3и Н4. Они образуют белковые тела или коры,состоящие из восьми молекул. Молекула ДНК комплексуется с белковыми корами, спирально накручиваясь на них-биспирали. Свободную от контакта с корами ДНК наз линкерной(связующая).Отрезок ДНК + белк кор = нуклеосома. Благодаря нуклеосомам в промоторных участках ДНК заблокированы области инициации (начала) транскрипции. Для того чтобы инициаторный комплекс возник, нуклеосомы должны быть «вытеснены» из соответствующих фрагментов ДНК. Образование хроматиновой фибриллы диаметром 30 нм (второй уровень компактизации) происходит с участием гистона Н1, который, связываясь с линкерной ДНК, скручивает нуклеосомную нить в спираль. На следующем петельно-доменном -укладка фибриллы диаметром 30 нм в петли. В этом процессе активная роль отводится негистоновым белкам. Основания петель «заякорены» в ядерном матриксе. Петля содержит от одного до нескольких генов(петельный домен). На следующем уровне компактизации «сложенные» фибриллы превращаются в метафазные хроматиды (хромосомы будущих дочерних клеток). Максимальная степень компактизации достигается на пятом уровне в структурах, известных как метафазные хромосомы с диаметром 1400 нм. Такая структура обеспечивает оптимальное решение задачи транспортировки генетического материала в дочерние клетки в анафазе митоза. 59. Динамика хромосомного материала в клеточном цикле.
60. Механизмы поддержания постоянства кариотипа поколений организмов и клеток. У организмов, размнож-ся бесполым путем, новое поколение появл в рез-те митоза, обеспеч таким образом сохранение постоянной стр-ры наследственного материала в ряду поколений клеток и орг-мов.
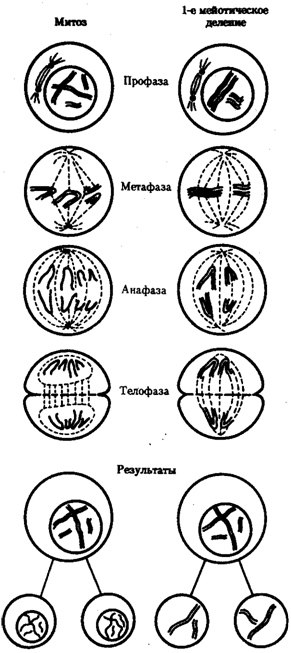
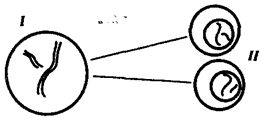 При половом размножении процесс осущ с помощью гамет, вступающих в оплодотворение. При оплодотворении наследственный материал двух родительских гамет сливается, образуя генотип организма нового поколения — зиготы. Чтобы потомки получили соответствующую программу для развития видовых и индивидуальных характеристик, они должны обладать кариотипом, которым располагало предыдущее поколение. Постоянство кариотипа в ряду поколений достигается уменьшением вдвое набора хромосом в гаметах, кот восстан до диплоидного при их оплодотворении: п + п = 2n. Образование гаплоидных гамет осущ в ходе гаметогенеза путем мейоза. При мейозе из клеток с диплоидным набором In образуются гаметы с гаплоидным набором хромосом п . Благодаря тому, что после однократного удвоения ДНК клетка делится дважды. В отличие от митоза в первом мейотическом делении в результате конъюгации гомологичные хромосомы объединяются в пары — биваленты. Последующее расхождение гомологов к разным полюсам веретена деления - гаплоидный набор хромосом: 2n4с → п2с. В ходе второго мейотического деления сестринские хроматиды каждой хромосомы, как и в митозе, распределяются между дочерними клетками с наследственным материалом пс. Рис. 3.71. Схема второго (эквационного) деления мейоза: I — кл, обр в рез-те 1-го мейотического деления=двунитчатых хром (n2c); II — кл, обр после 2-го деления мейоза и несущ гаплоидный набор однонитчатых хром (nc) Благодаря особенностям мейоза образуются клетки, несущ полноценный геном, в кот каждая группа сцепления представлена в единственном экземпляре (гаплоидный набор хромосом). Сперматозоиды, проникая в яйцеклетку, вводят в нее свой ядерный наследственный материал, заключенный в гаплоидном наборе хромосом. Ядра гамет сливаются = диплоидное ядро зиготы, в кот каждая группа сцепления представлена в двойном экземпляре — отцовской и материнской хромосомами. Таким образом, мейоз и последующее оплодотворение обеспечивают сохранение у нового поколения организмов диплоидного кариотипа, присущего всем особям данного вида. 61. Жизненный цикл клетки и его периоды. 1. Пресинтетический или постмитотический (G1) период наступает сразу же после митотического деления клетки и характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему клетка достигает нормальных размеров и восстанавливает необходимый набор органелл. G1 -период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые "запускающие" белки, или активаторы S-периода. Они обеспечивают достижение клеткой определенного порога (точки R - рестрикции или ограничения), после которого она вступает в S-период. Контроль, осуществляемый на уровне точки R (при переходе из G1 в S), ограничивает возможность нерегулируемого размножения клеток. Проходя эту точку, клетка переключается на последующую регуляцию внутренними факторами клеточного цикла, которая обеспечивает закономерное завершение ее деления. Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0) для того, чтобы (в зависимости от причин остановки): -дифференцироваться и выполнять свои специфической функции -выжить в условиях недостаточности питательных веществ или факторов роста -осуществить репарацию поврежденной ДНК. Клетки одних тканей при соответствующей стимуляции вновь способны возвращаться из периода (G0) в клеточный цикл, других - утрачивают эту способность по мере дифференцировки | ||||||||||||||||||||||||||||||||