1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
2. Синтетический (S) характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов. 3. Постсинтетический или премитотический (G2) период следует за S-периодом и продолжается вплоть до митоза (часто обозначаемого буквой М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (в частности, тубулин), необходимые для процесса деления. Длительность G2-периода составляет 2-4 часа. Контроль вступления клетки в митоз осуществляется двумя специальными факторами с противоположно направленными эффектами. Митоз тормозится до момента завершения репликации ДНК М-задерживающим фактором и индуцируется М-стимулирующим фактором. Действие последнего проявляется лишь в присутствии других белков - циклинов (синтезируются на протяжении всего цикла и распадаются в середине митоза). 62. Нарушения клеточного цикла. Амитоз. Эндомитоз. Политения. Эндомитоз (от греч. endon -внутри и mitos - нить) - вариант митоза, при котором происходит удвоение числа хромосом внутри ядерной оболочки без ее разрушения и образования веретена деления. При повторных эндомитозах число хромосом в ядре может значительно увеличиваться при соответствующем кратном двум нарастании содержания в нем ДНК - полиплоидии (от греч. poly - много и ploon - складываю) и увеличении объема ядра. Полиплоидия может явиться также результатом неоконченных обычных митозов. Основной смысл развития полиплоидии заключается в усилении функциональной активности клетки. Сходный результат достигается при образовании двуядерных клеток вследствие митотического деления, не сопровождающегося цитотомией. При последующем митотическом делении такой двуядерной клетки хромосомные наборы ядер объединяются в метафазе, приводя к образованию двух дочерних полиплоидных клеток. Наличие полиплоидных - тетра- (4n) и октаплоидных (8n) клеток - нормальное явление в печени, эпителии мочевого пузыря, клетках концевых отделов поджелудочной и слюнных желез. Мегакариоциты (гигантские клетки костного мозга) начинают формировать кровяные пластинки лишь достигнув определенного уровня полиплоидии (16-32n) в результате нескольких эндомитозов. Другой тип полиплоидизации-политения. При политении происходит многократная репликация нитчатых интерфазных хромосом без последующего расхождения, митоза. Клетки с политенными хромосомами никогда не делятся. В результате возникают продольные многонитчатые структуры, или политенные хромосомы. Амитоз, или прямое деление, — это деление интерфазного ядра путем перетяжки. При амитозе веретено деления не образуется, отсутствуют все четыре фазы, свойственные митозу, спирализации хромосом не происходит, поэтому в световой микроскоп они не видны. Такое деление встречается в тканях растущего клубня картофеля, в клетках эндосперма, стенок завязи пестика, печени и хряща, роговицы глаза. Распределение клеточных органоидов, как и ДНК, осуществляется произвольно. Амитоз в отличие от митоза является самым экономичным способом деления, так как энергетические затраты при этом незначительны. 63. Бесполое размножение и его формы. Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение. Деление-способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить: -простое бинарное деление (прокариоты) -митотическое бинарное деление (простейшие, одноклеточные водоросли)- -множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время шизогонии сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов. Почкование-способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи. Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы). Фрагментация-способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации. Полиэмбриония-способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы). Вегетативное размножение-способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение). 64. Митоз – цитологическая основа бесполого размножения. Митоз (от греч. mitos - нить) является универсальным механизмом деления клеток. Митоз следует за G2-периодом и завершает клеточный цикл. Он длится 1-3 часа и обеспечивает равномерное распределение генетического материала в дочерние клетки. Митоз включает 4 основные фазы профазу, метафазу, анафазу и телофазу. Профаза. Эта стадия характеризуется постепенной конденсацией и спирализацией хромосом. Под микроскопом становятся различимы хромосомы в виде нитевидных структур. Видно, что каждая хромосома состоит из двух копий-дочерних хроматид, соединенных центромерой. Другим важным событием профазы является постепенное исчезновение ядрышка, содержимое которого распределяется по ядру. К полюсам клетки расходятся дочерние центриоли, и между ними формируется митотическое веретено. Митотическое веретено-это биполярная структура, которая состоит из микротрубочек. Метафаза. На этой стадии хромосомы, прикрепленные центромерами к нитям веретена, собираются на экваторе-плоскости, расположенной посередине между полюсами веретена. Образовавшаяся метафазная пластинка представляет собой наиболее характерную особенность метафазы. Хромосомы максимально спирализованы. Анафаза. На этой стадии каждая центромера делится пополам. Хроматиды становятся самостоятельными хромосомами, и две дочерние ценромеры устремляются к противоположным полюсам, увлекая за собой по одной из двух дочерних хромосом. Телофаза. Два набора дочерних хромосом группируются у противоположных полюсов веретена. Происходит деспирализация хромосом. Вокруг каждого набора хромосом образуется ядерная оболочка, вновь возникает ядрышко. Цитокинез. Разделение цитоплазмы на две обособляющиеся части. Механическим аппаратом цитокинеза является сократимое кольцо микрофиламентов, которые состоят из актина. Митотическое веретено и сократимое кольцо располагаются перпендикулярно друг другу, и веретено оказывается внутри сократимого кольца. Борозда деления разделяет плоскость митоза и образуется две генетически равноценные клетки. Цитокинез начинается в телофазе. 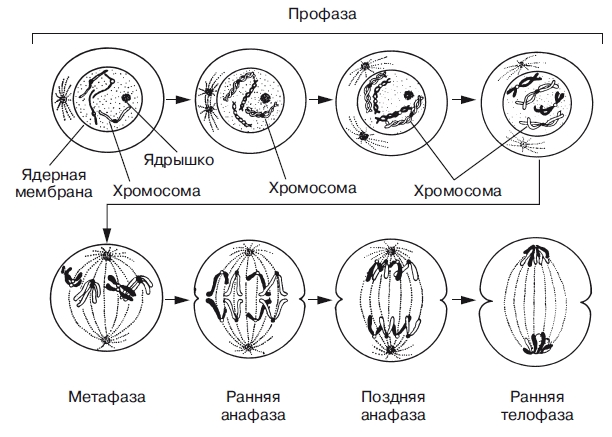 65. Половое размножение. Регулярные и нерегулярные формы. Половое размножение встречается в основном у высших организмов. Оно обеспечивает значительное генетическое разнообразие и, следовательно, большую фенотипическую изменчивость потомства; организмы получают большие эволюционные возможности, возникает материал для естественного отбора. Помимо полового размножения, существует половой процесс. Суть его в том, что обмен генетической информацией между особями происходит, но без увеличения числа особей. Формированию гамет у многоклеточных предшествует мейоз. Половой процесс состоит в объединении наследственного материала от двух разных источников (родителей). При половом размножении потомство генетически отличается от своих родителей, так как между родителями происходит обмен генетической информацией. Основой полового размножения является мейоз. Родителями являются две особи – мужская и женская, они вырабатывают разные половые клетки. В этом проявляется половой диморфизм, который отражает различие задач, выполняемых при половом размножении мужским и женским организмами. Половое размножение осуществляется через гаметы – половые клетки, имеющие гаплоидный набор хромосом и вырабатывающиеся в родительских организмах. Слияние родительских клеток приводит к образованию зиготы, из которой в дальнейшем образуется организм-потомок. Половые клетки образуются в гонадах – половых железах (в яичниках у самок и семенниках у самцов). Процесс образования половых клеток называется гаметогенезом (овогенезом у самок и сперматогенезом у самцов). Если мужские и женские гаметы образуются в организме одной особи, то ее называют гермафродитной. Гермафродитизм бывает истинный (особь имеет гонады обоих полов) и ложный гермафродитизм (особь имеет половые железы одного типа – мужского или женского, а наружные половые органы и вторичные половые признаки обоих полов). Партеногенез (девственное размножение) Виды партеногенеза: 1.облигатный (обязательный) партеногенез. Встречается в популяциях, состоящих исключительно из особей женского пола. При этом вероятность встречи разнополых особей минимальна 2.циклический (сезонный) партеногенез (у тлей, дафний, коловраток). Встречается в популяциях, которые исторически вымирали в больших количествах в определенное время года. У этих видов партеногенез сочетается с половым размножением. При этом в летнее время существуют только самки, которые откладывают два вида яиц – крупные и мелкие. Из крупных яиц партеногенетически появляются самки, а из мелких – самцы, которые оплодотворяют яйца, лежащие зимой на дне. Из них появляются исключительно самки; 3.факультативный (необязательный) партеногенез. Встречается у общественных насекомых (ос, пчел, муравьев). В популяции пчел из оплодотворенных яиц выходят самки (рабочие пчелы и царицы), из неоплодотворенных – самцы (трутни). Гиногенез (у костистых рыб и некоторых земноводных). Сперматозоид проникает в яйцеклетку и лишь стимулирует ее развитие. Ядро сперматозоида при этом с ядром яйцеклетки не сливается и погибает, а источником наследственного материала для развития потомка служит ДНК ядра яйцеклетки. Андрогенез. В развитии зародыша участвует мужское ядро, привнесенное в яйцеклетку, а ядро яйцеклетки при этом гибнет. Яйцеклетка дает лишь питательные вещества своей цитоплазмы. Полиэмбриония. Зигота (эмбрион) делится на несколько частей бесполым способом, каждая из которых развивается в самостоятельный организм. Встречается у насекомых (наездников), броненосцев. У броненосцев клеточный материал первоначально одного зародыша на стадии бластулы равномерно разделяется между 4–8 зародышами, каждый из которых в дальнейшем дает полноценную особь. К этой категории явлений можно отнести появление однояйцовых близнецов у человека. У одноклеточных организмов выделяют две формы полового размножения – копуляцию и конъюгацию. При конъюгации (например, у инфузорий) специальные половые клетки не образуются. При этом процессе не происходит увеличения количества особей, поэтому говорят о половом процессе, а не о половом размножении. Однако происходит обмен (рекомбинация) наследственной информацией, поэтому потомки генетически отличаются от своих родителей. При копуляции (у простейших) происходят образование половых элементов и их попарное слияние. При этом две особи приобретают половые различия и полностью сливаются, образуя зиготу. Происходят объединение и рекомбинация наследственного материала, поэтому особи генетически отличны от родительских. 66. Цитологические основы полового размножения. Мейоз, как специфический процесс при формировании половых клеток. Мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным. Во втором делении число хромосом не изменяется; такое деление называют эквационным. Интерфаза Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с. Первое деление мейоза (редукционное деление, или мейоз I) Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды). Профаза 1 состоит из ряда стадий: Лептотена. Хромосомы видны в световой микроскоп в виде клубка тонких нитей. Раннюю лептотену, когда нити хромосом видны еще очень плохо, называют пролептотена. Зиготена. Происходит конъюгация гомологичных хромосом. Гомологичные хромосомы – это хромосомы, сходные между собой в морфологическом и генетическом отношении. При конъюгации образуются биваленты. Каждый бивалент – это относительно устойчивый комплекс из одной пары гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды. Пахитена. Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК. Завершается кроссинговер. Диплотена. Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы. Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра. Метафаза I. Ядерная оболочка разрушается (фрагментируется). Формируется веретено деления. Далее происходит метакинез – биваленты перемещаются в экваториальную плоскость клетки. Анафаза I.Гомологичные хромосомы, входящие в состав каждого бивалента, разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит. Процесс распределения хромосом по дочерним клеткам называется сегрегация хромосом. Телофаза I.Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются два гаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с. В большинстве случаев (но не всегда) телофаза I сопровождается цитокинезом. Интеркинез-это короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы тем, что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей. Второе деление мейоза (эквационное деление, или мейоз II) В ходе второго деления мейоза уменьшения числа хромосом не происходит. Сущность эквационного деления заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида). Профаза II. Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. Метафаза II.В каждой из дочерних клеток формируется веретено деления. Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток независимо друг от друга. Эти экваториальные плоскости могут лежать в одной плоскости, могут быть параллельны друг другу или взаимно перпендикулярны. Анафаза II.Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы в составе анафазных групп перемещаются к полюсам клеток. Телофаза II.Однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с. 67. Гаметогенез и его этапы. Сравнение овогенеза и сперматогенеза.  Гаметогенез — это процесс образования половых клеток. Протекает он в половых железах (в яичниках у самок и в семенниках у самцов). Гаметогенез в организме женской особи сводится к образованию женских половых клеток (яйцеклеток) и носит название овогенеза. У особей мужского пола возникают мужские половые клетки (сперматозоиды), процесс образования которых называется сперматогенезом. Стадии гаметогенеза 1. Стадия размножения. Клетки, из которых в последующем образуются мужские и женские гаметы, называются сперматогониями и овогониями соответственно. Они несут диплоидный набор хромосом 2n2c. На этой стадии первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает. Сперматогонии размножаются в течение всего репродуктивного периода в мужском организме. Размножение овогоний происходит главным образом в эмбриональном периоде. 2. Стадия роста. Kлетки увеличиваются в размерах(за счет увеличения объема цитоплазмы) и превращаются в сперматоциты и овоциты I порядка (последние достигают особенно больших размеров в связи с накоплением питательных веществ в виде желтка и белковых гранул). Эта стадия соответствует интерфазе I мейоза. Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c. 3. Стадия созревания. Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз. После первого деления (мейоза I) образуются сперматоциты и овоциты II порядка (с генетической формулой n2c), после второго деления (мейоза II) — сперматиды и зрелые яйцеклетки (с формулой nc) с тремя редукционными тельцами, которые погибают и в процессе размножения не участвуют. Таким образом, в результате стадии созревания один сперматоцит I порядка (с формулой 2n4c) дает четыре сперматиды (с формулой nc), а один овоцит I порядка (с формулой 2n4c) образует одну зрелую яйцеклетку (с формулой nc) и три редукционных тельца. 4. Стадия формирования, или спермиогенеза (только при сперматогенезе). В результате этого процесса каждая незрелая сперматида превращается в зрелый сперматозоид (с формулой nc), приобретая все структуры, ему свойственные. Ядро сперматиды уплотняется, происходит сверхспирализация хромосом, которые становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, формируя акросому. К другому полюсу ядра устремляются центриоли, причем одна из них принимает участие в формировании жгутика. Вокруг жгутика спирально закручивается одна митохондрия. Почти вся цитоплазма сперматиды отторгается, поэтому головка сперматозоида ее почти не содержит. Отмеченные выше различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество. Центральное событие в процессе гаметогенеза — редукция диплоидного набора хромосом (в ходе мейоза) и формирование гаплоидных гамет. 68. Закономерности сперматогенеза у млекопитающих и человека.(схема 67) Последовательные стадии развития сперматозоидов получили название сперматогенеза. 1. Стадия размножения. Клетки, из которых в последующем образуются мужские гаметы, называются сперматогониями. Они несут диплоидный набор хромосом 2n2c. На этой стадии первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает. Сперматогонии размножаются в течение всего репродуктивного периода в мужском организме. 2. Стадия роста. Kлетки увеличиваются в размерах (за счет увеличения объема цитоплазмы) и превращаются в сперматоциты I порядка Эта стадия соответствует интерфазе I мейоза. Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c. 3. Стадия созревания. Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз. После первого деления (мейоза I) образуются сперматоциты II порядка (с генетической формулой n2c), после второго деления (мейоза II) — сперматиды (с формулой nc) . Таким образом, в результате стадии созревания один сперматоцит I порядка (с формулой 2n4c) дает четыре сперматиды (с формулой nc), 4. Стадия формирования, или спермиогенеза (только при сперматогенезе). Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферических неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид (с формулой nc). Ядро сперматиды уплотняется, происходит сверхспирализация хромосом, которые становятся функционально инертными. Комплекс Гольджи перемещается к одному из полюсов ядра, формируя акросому. К другому полюсу ядра устремляются центриоли, причем одна из них принимает участие в формировании жгутика. Вокруг жгутика спирально закручивается одна митохондрия. Почти вся цитоплазма сперматиды отторгается, поэтому головка сперматозоида ее почти не содержит. Каждый сперматозоид человека состоит из трех отделов — головки, средней части и хвоста. В головке сперматозоида располагается ядро. В нем содержится гаплоидный набор хромосом. Головка снабжена акросомой, которая содержит литические ферменты, необходимые сперматозоиду для вступления в яйцеклетку. В головке локализуется также две центриоли — проксимальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются базальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью. Различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество. 69. Закономерности овогенеза у млекопитающих и человека. (Схема.67) Гаметогенез в организме женской особи сводится к образованию женских половых клеток (яйцеклеток) и носит название овогенеза. Он протекает он в половых железах (в яичниках у самок). Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и обеспечение питательных потребностей зиготы Стадии овогенеза 1.Стадия размножения. Первичные половые клетки многократно делятся митозом, в результате чего их количество существенно возрастает. Размножение оогоний происходит главным образом в эмбриональном периоде. 2. Стадия роста. Kлетки увеличиваются в размерах (за счет увеличения объема цитоплазмы) и превращаются в овоциты I порядка. Эта стадия соответствует интерфазе I мейоза. Важное событие этого периода — репликация молекул ДНК при неизменном количестве хромосом. Они приобретают двунитчатую структуру: генетическая формула клеток в этот период выглядит как 2n4c. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро. 3. Стадия созревания. Происходят два последовательных деления — редукционное (мейоз I) и эквационное (мейоз II), которые вместе составляют мейоз. Ко времени рождения девочки все первичные ооциты уже находятся в профазе первого мейотического деления. Ооцит в состоянии покоя окружен одним слоем уплощенных фолликулярных клеток и образует вместе с ними примордиальный фолликул. Ооциты, не окруженные фолликулами, дегенерируют. Некоторые из фолликулов периодически начинают развитие-фолликулогенез. Сначала клетки фолликула делятся, образуя многослойную оболочку ооцита; сам ооцит растет, вокруг него образуется прозрачная многослойная зона-формируется развивающийся фолликул. Затем в некоторых фолликулах образуется зона-формируется развивающийся фолликул. Затем в некоторых фолликулах образуется наполненная жидкостью полость-антриум, и развивающийся фолликул превращается в антральный. Внутри фолликула ооцит постепенно окружается клеточной массой яйценосного бугорка. До достижении половой зрелости процессы развития фолликулов завершаются дегенерацией. С наступлением половой зрелости также одновременно развиваются многие фолликулы, однако лишь некоторые высвобождают находящиеся в них ооциты в процессе овуляции. Большая часть фолликулов дегенерирует на той или иной стадии. Продолжительность роста фолликула составляет приблизительно 6 месяцев. В соответствии с менструальным циклом раз в месяц происходит повышение уровня лютеинизирующего гормона, выделяемого гипофизом, что побуждает примерно 20 антральных фолликулов к ускоренному росту. Однако завершает созревание и претерпевает овуляцию лишь один из фолликулов. Ооцит первого порядка, находящийся в этом фолликуле, завершает первое деление мейоза, образуя полярное тельце и превращаясь в ооцит второго порядка. Ооцт второго порядка, развитие которого останавливается на стадии метафазы второго деления мейоза, освобождается вместе с полярным тельцем и часть окружающих его клеток в момент разрыва фолликула на поверхности яичника. Ооцит второго порядка завершает второе мейотическое деление только в том случае, если он будет оплодотворен. После овуляции опустевший фолликул превращается в эндокринное образование-желтое тело. Оно секретирует прогестерог, готовя матку к приему оплодотворенного ооцита. Если оплодотворение не происходит, желтое тело деградирует, а слизистая оболочка матки отслаивается и выводится из организма во время менструации. Отличительные особенности Питательный материал (желток) первичного ооцита не распределяется поровну между четырьмя клетками.. Основное количество желтка сохраняется в одной большой клетке. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные ооциты, но они не становятся половыми клетками. Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характерные для соматических клеток. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. Различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется на стадии ооцита и представляет собой поверхностный слой ооцита. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. Различия в ходе овогенеза и сперматогенеза имеют определенный биологический смысл, связанный с разным функциональным назначением мужских и женских гамет (помимо переноса генетической информации). Накопление в цитоплазме яйцеклетки большого количества запасных питательных веществ необходимо, так как на этой «базе» осуществляется развитие дочернего организма из оплодотворенного яйца. Неравномерное клеточное деление при овогенезе и обеспечивает формирование крупной яйцеклетки. Функция же сперматозоидов заключается в отыскании яйцеклетки, проникновении в нее и доставке своего хромосомного набора. Их существование кратковременно, а поэтому нет необходимости в запасании большого количества веществ в цитоплазме. А поскольку сперматозоиды в массе гибнут в процессе поиска яйцеклетки, их образуется огромное количество. 70. Оплодотворение, его формы и биологическая функция. Моно- и полиспермия. Оплодотворение-это процесс объединения мужской и женской гамет, который приводит к формированию зиготы и последующему развитию нового организма. В процессе оплодотворения происходит установление диплоидного набора хромосом в зиготе, что определяет выдающееся биологическое значение этого процесса. В зависимости от видовой принадлежности организмов у животных, размножающихся половым путем, различают наружное и внутреннее оплодотворение. Наружное оплодотворение происходит в окружающей среде, в которую поступают мужские и женские половые клетки. Например, наружным является оплодотворение у рыб. Выделяемые ими мужские (молока) и женские (икра) половые клетки поступают в воду, где и происходит их «встреча» и объединение. Внутреннее оплодотворение обеспечивается переносом сперматозоидов из мужского организма в женский в результате полового акта. Такое оплодотворение встречается у млекопитающих, причем центральным моментом здесь является исход встречи между половыми клетками. Считают, что в яйцеклетку этих животных проникает ядерное содержимое лишь одного сперматозоида. Что касается цитоплазмы сперматозоида, то у одних животных она поступает в яйцеклетку в небольшом количестве, у других совсем не поступает в яйцеклетку. У человека оплодотворение происходит в верхней части фаллопиевой трубы, причем в оплодотворении, как и у других млекопитающих, участвует лишь один сперматозоид, ядерное содержимое которого поступает в яйцеклетку. Иногда в фаллопиевой трубе может оказаться не одна, а две или более яйцеклеток, в результате чего возможно рождение двоен, троен и т. д. В результате оплодотворения в оплодотворенной яйцеклетке восстанавливается диплоидный набор хромосом. Яйцеклетки способны к оплодотворению в течение примерно 24 часов после овуляции, тогда как оплодотворяющая способность сперматозоидов сохраняется до 48 часов. В зависимости от количества особей, принимающих участие в половом размножении, различают: -перекрестное оплодотворение-оплодотворение, в котором принимают участие гаметы, образованные разными организмами -самооплодотворение-оплодотворение, при котором сливаются гаметы, образованные одним и тем же организмом (ленточные черви). Существует два типа оплодотворения: моно- и полиспермия. При моноспермии благодаря особым механизмам в яйцеклетку проникает лишь один сперматозоид. Этот вид оплодотворения широко распространен у животных с наружным осеменением, хотя свойствен также и млекопитающим. При полиспермии в яйцеклетку проникает до нескольких десятков сперматозоидов, однако ядро только одного из них соединяется с женским пронуклеусом, остальные посредством специальных механизмов исключаются из развития. Этот тип оплодотворения присущ животным с внутренним осеменением (членистоногие, моллюски, хордовые). Оболочка яйцеклетки насекомых и многих рыб содержит отверстия, через которые сперматозоиды проникают в яйцеклетку. У млекопитающих такие отверстия отсутствуют, и, чтобы достигнуть ооплазмы, сперматозоиды должны пройти через толщу оболочек. Для этого у сперматозоида имеются специальные органеллы-акросомы, располагающиеся на вершине его головки. Погрузившись в ооплазму, головка сперматозоида ориентируется в глубь яйцеклетки и претерпевает сложные изменения. При этом его ядро постепенно преобразуется в мужской пронуклеус. Эти преобразования идут одновременно с формированием женского пронуклеуса. Мужской и женский пронуклеусы сближаются в центре яйцеклетки и затем сливаются, образуя единое ядро зиготы. На этом завершается процесс оплодотворения. Образовавшаяся после оплодотворения зигота постепенно продвигается по направлению к матке и выходит в нее через несколько суток. 71. Морфологические и функциональные особенности зрелых гамет млекопитающих и человека Гаметы - репродуктивные клетки, имеющ гаплоидный (одинарный) набор хром и уч в гаметном, половом размножении. При слиянии двух гамет обр зигота, развив в особь (или группу особей) с наследственными признаками обоих родительских организмов, продуцировших гаметы. У некоторых видов возможно и развитие в организм одиночной гаметы (неоплодотворённой яйцеклетки) — партеногенез. Морфология гамет и типы гаметогамии Продуцируемые гаметы могут отличаться как по хромосомному набору (при гетерогаметности вида), величине и подвижности (способности к самостоятельному передвижению). Если сливающиеся гаметы морфолог не отличаются друг от друга величиной, строением и хромосомным набором-это изогаметы(бесполые гам). Они подвижны, могут нести жгутики или быть амёбовидными. Изогамия типична для многих водорослей. Анизогамия (гетерогамия) Гаметы, способные к слиянию, различаются по размерам, подвижные микрогаметы несут жгутики, макрогаметы могут быть как подвижны (многие водоросли), так и неподвижны (лишённые жгутиков макрогаметы многих протистов). Оогамия Сперматозоид и яйцеклетка. Способные к слиянию гаметы одного биологического вида резко различаются по размерам и подвижности на два типа: мужскиегаметымалого размера и крупные неподвижные женскиегаметы— яйцеклетки. Различие размера гамет обусловлено тем, что яйцеклетки содержат запас питательных веществ, достаточный для обеспечения нескольких первых делений зиготы при её развитии в зародыш. Мужские гаметы — сперматозоиды животных и многих растений подвижны и обычно несут один или несколько жгутиков, исключением являются лишённные жгутиков мужские гаметы семенных растений — спермии, кот дост-ся к яйцеклетке при прорастании пыльцевой трубки, безжгутиковые сперматозоиды (спермии) нематод и членистоногих. Хотя сперматозоиды несут митох, при оогамии от мужской гаметы к зиготе переходит ядерная ДНК, мтх ДНК наследуется зиготой только от яйцеклетки. 72. Клеточные факторы иммунной системы. -лимфоциты(Т, B, NK) -моноциты -макрофаги -нейтрофилы -эозинофилы -базофилы(тучные) клетки Клеточн факторы врожденного иммунитета: Нейтрофилы и Макрофаги(моноциты)-«профессиональные» фагоциты. Для уничтожения: 1. Кислороднонезависимый мех-м(гидролитические ферм) 2. Кислородзависимый(«кислородн» взрыв с обр н2о2 и аниона о2-, гипохлорида-для киллинга) 3. NO (и производные) после дополн активац с помощью Т-хелперов. Продукция и секреция: +Вещ-ва для уничтожения прод вне клетки +Сигнальные вещ-ва (цитокинов и тд…) МОНОЦИТЫ и МАКРОФАГИ – Антигенпредставл кл (АПК) Представляют антигены-презентац антигенов в компл с МНС (главн комп гистосовместим) Т-клетками. ДЕНДРИТНЫЕ кл-с отростками. - захват, затем процессинг антигена - презентац антигенов Т-лимфоцитам (гл АПК) - костимулирующие сигналы Т-лимф - др. функц-регуляц созревания Т-лимф в тимусе БАЗОФИЛЫ(тучн кл) – Синтез и секреция медиаторов воспаления(гистамин, простагландин, гепарин…) Вызыв аллергич реакц, несут на поверхн рецепторы к иммуноглобулинам Е и Fe ЭОЗИНОФИЛЫ – антителзависимое цитотоксическое вещ-во(действ в отношении личинок гельминтов) Уч в аллергич реакции, несут рецепторы к иммуноглобулинам Е и Fe NK-клетки (норм киллеры)-лимфоциты. Уничтожают кл опухолей и инфицированные вирусами. Цитотоксич действие-без апоптоза в отлич от Т и В лимфоцитов. Не имеют антиген спецефич рецепторов. Убивают кл, лишившиеся MHCl. Проявл антителзависимую кл цитотоксичность (связ «хвост» антитела с помощбю Fe рец и убивают кл с кот это антитело взаимодействует). 73. Т-лимфоциты – клеточные факторы приобретенного клеточного иммунитета. Термин клеточный иммунитет используется для описания такого противоинфекционного или противоопухолевого иммунного ответа, в котором антителам принадлежит не ведущая, а вспомогательная роль. Основу клеточного иммунитета составляют лимфоциты, которые для своего созревания переселяются из костного мозга в другой центральный орган лимфоидной системы - тимус (вилочковая железа). Эта ветвь лимфоцитов получила название тимус-зависимые, или Т-лимфоциты. Т-лимфоциты выполняют следующие функции: – являются основными эффекторами клеточного иммунитета (эти клетки опосредуют клеточные цитотоксические реакции, а также реакции ГЗТ); – являются регуляторами воспаления, иммунных реакций и гемопоэза; – участвуют в процессах репаративной и физиологической регенерации различных тканей. В организме человека Т-лимфоциты многократно покидают лимфоидные органы, попадая сначала в лимфу, затем в кровь, а из крови снова возвращаются в органы. За свою жизнь лимфоцит может проходить более 100 километров. Благодаря интенсивной циркуляции, лимфоциты, когда в них возникает потребность, быстро появляются в "горячих точках". В тимусе формируются разные виды Т-клеток. 1.некоторые Т-лимфоциты обладают способностью разрушать клетки, содержащие чужеродный антиген, их назвали цитотоксическими или "киллерами". 2.другая разновидность лимфоцитов - Т-хелперы- первыми распознают чужеродные вещества. Т-хелперы не способны вырабатывать антитела и убивать клетки-мишени, но, распознавая чужеродный антиген, они реагируют на него выработкой различных факторов, которые необходимы для размножения и созревания В-клеток и Т-киллеров. Именно Т-хелперы распознают антигены и влияют на деятельность других типов Т-клеток, оказывают помощь В-клеткам в образовании антител. По их командам иммунная система направляет Т-лимфоцитов-киллеров, задача которых убивать зараженные клетки. 3.существуют еще Т-супрессоры, которые подавляют активность иммунного ответа, когда необходимость в нем отпадает. Если иммунные клетки будут продолжать работать, то будут поражаться собственные здоровые клетки организма, что приведет к развитию различных болезней (их называют аутоиммунными). 74. В-лимфоциты – клеточные факторы приобретенного гуморального иммунитета. Приобретенный иммунитет: • Т и В лимфоц • MHC (у чел HLA) • Антитела • Клеточный и Гуморальный иммунный ответ В-лимфоциты-гуморальный иммун. Распознание нерасщепл антигенов. Наивные В-лимф. Плазматические В-кл памяти-вторичный иммунитет. Отличия Т- и В-лимфоцитов: - по месту антигеннезависим дифференц: Т-лимфоциты в тимусе, В-лимфоциты в лимфоидной ткани других органов. - по выполняемым функциям: Т-лимфоциты обеспечивают преимущественно клеточный иммунитет, В-лимфоциты – гуморальный иммунитет. Функционально Т- и В-клетки делят на субпопуляции. Среди Т-лимфоцитов выделяют Тх (хелперы) – активируют эффекторные клетки, Тк (киллеры) – эффекторные цитотоксические клетки, Тс (супрессоры) – подавляют иммунный ответ, Т-лимфоциты памяти. В-лимфоциты дифференцируются в плазматические клетки, вырабатывающие иммуноглобулины (антитела) и в клетки памяти, несущие информацию о встрече с каким- либо антигеном. - по содержанию в крови: Т-лимфоцитов – 70-80%, В-лимфоцитов — 10-20%. Функция лимфоцитов: 1) участие и обеспечение иммунных реакций.  В-лимфоциты проходят предварительную обработку в печени и костном мозге. Существуют два важных различия между В- и Т-лимфоцитами. - Во-первых, В-лимфоциты активно секретируют реактивные агенты, называемые антителами, в отличие от Т-лимфоцитов, реагирующих с антигеном непосредственно. Антитела — это крупные белковые молекулы, способные соединяться с антигенной субстанцией и разрушать ее. - Во-вторых, разнообразие В-лимфоцитов выражено больше, чем у Т-лимфоцитов, т.е. формируются миллионы типов В-лимфоцитарных антител с разными специфическими реактивностями.  |