1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
23. Принципы и механизмы преобразования сигналов в клетке. -Изменения конформации белков -Изменение активности путем фосфо и дефосфорилирования белков (фосф к-ты из АТФ) -Преобразование сигнала с уч. G-белков -Обравзование вторичных посрелников Ферменты: 1)Киназы-фосфорилировае. 2)Фосфотазы-дефосфорилирование Каталитические рецепторы. Белки-рецепторы.(Инсулин, и многие фкторы роста) У кот домен облад ферментативной(киназной)СПОСОБНОСТЬЮ АКТИВАЦИИ БЕЛКОВ. G-белки – разновидность GTPазы, стр и активность кот зависит от нахождения ГТФ или ГДФ-связном состоянии. Участвуют в передаче неклеточных сигналов от рецепторов в клетку (сопрягая активацию рецепторов в работой ионных каналов или ферментов) РОЛЬ G-белков: (Гилман и Мартин-открытие) •Усиление сигнала •Возможность регудяции активности белков-мишений.(G5-стимулирующ.G1-ингибирующ белок) (Активац калиевого канала с уч G-белка) В системе ц-АМФ(Аденилатциклазная), внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке. Участие аденилатциклазной системы в регуляции экспрессии генов. Многие белковые гормоны: глюкагон, вазопрессин, паратгормон и др. могут не только изменять скорости реакций путем фосфорилирования ферментов, но и 24. Межклеточные соединения (контакты). Адгезивные соединения. Адгезивные (механические) контакты-это клеточные контакты, обеспечивающие механическую связь между клетками. Способствуют равномерному распределению механической нагрузки на весь пласт клеток, защищая мембраны от деформации. Соединяют плазматические мембраны соседних клеток и укрепляют их через связь с фибриллами цитоскелета. В их формирование обязательно участвует два типа белков: трансмембранные связующие и внутриклеточные. К механическим контактам относят точечные десмосомы, полудесмосомы и опоясывающие десмосомы. Точечные разбросаны по поверхности контактирующих клеток. Полудесмосомы сходны по строению с десмосомами, но представляют собой соединение клеток с межклеточными структурами. Опоясывающие десмосомы располагаются сплошным пластом на всем протяжении контактирующих поверхностей. В формировании контактов этого типа принимают участие сократимые белки субмембранного комплекса. Характерный пример механического контакта-точечная десмосома. В такой десмосоме плазматические мембраны соседних клеток находятся на расстоянии 20-35 нм, между ними видна зона повышенной плотности в виде волокнистого матрикса (место взаимодействия трансмембранных гликопротеинов, которые при участии ионов кальция обеспечивают механическое соединение мембран двух соседних клеток). С внутренней стороны клеток к плазматической мембране прилегают плотные пластинки, состоящие из белка десмоплакина. С десмоплакином связаны промежуточные филаменты, обеспечивающие равномерное распределение нагрузки на весь клеточный пласт. Механические контакты чаще всего встречаются в тканях, испытывающих большие нагрузки. Они поддерживают целостность ткани, придают ткани прочность. 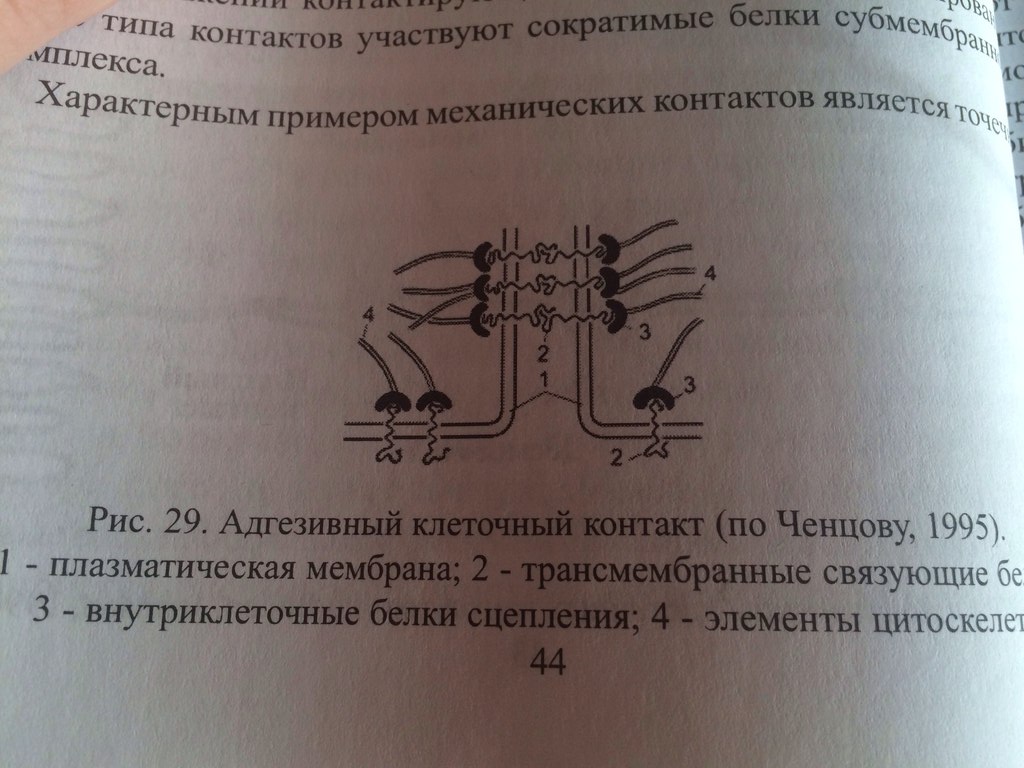 25. Межклеточные соединения. Проводящие (химические) контакты. 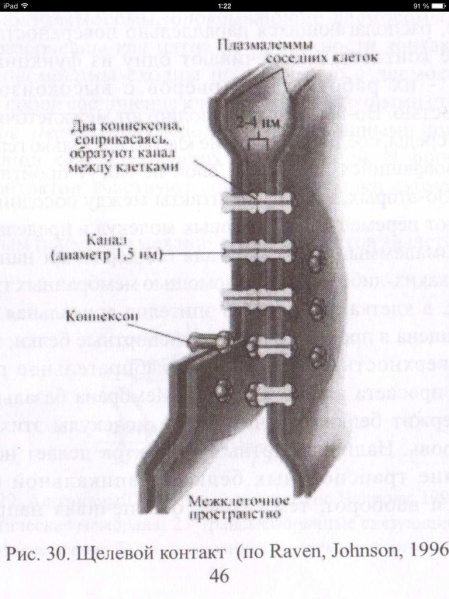 Химические контакты являются самым распространенным типом клеточных контактов в животных клетках. Они позволяют клеткам обмениваться небольшими водорастворимыми молекулами (неорганические ионы, сахара, аминокислоты…). Такой обмен обеспечивает электрическое и метаболическое сопряжение клеток. Делают возможной метаболическую кооперацию клеток (малые молекулы, производимые определенными клетками могут использоваться и остальными клетками). Разновидностями химических контактов: щелевые контакты, плазмодесмы растительных клеток и синапсы. Щелевые контакты построены из трансмембранных белков, которые формируют коннексоны (комплекс из 6 глобул интегрального белка коннексина, образующие ионный канал, контактирующий с аналогичным каналом соседней клетки). Плазмодесмы растительных клеток не являются собственно межклеточными контактами, а представляют собой места объединения клеток в надклеточную систему. Это миниатюрные каналы слияния плазматических мембран соседних клеток, происходит сообщение цитоплазмы клеток. Синапс-контакт между аксоном нейрона и любыми возбудимыми образованиями. Обеспечивает передачу сигнала с помощью молекул химических веществ. Пример: химический синапс между двумя нервными клетками. Аксон, подходя к другому нейрону, образует синаптическую бляшку. Непосредственного контакта между мембранами нейронов нет. Между ними-синаптическая щель (10-50 мкм). Мембрана синаптической бляшки-пресинаптическая, мембрана за синаптической щелью-постсинаптическая. В синаптической бляшке имеются синаптические пузырьки, содержащие нейромедиатор. Наружная поверхность мембраны нейрона заряжена положительно, внутренняя-отрицательно. Таки образом, существует разность потенциалов. Когда нервный импульс достигает окончания аксона, происходит деполяризация его мембраны. Она вызывает временное открывание кальциевых каналов мембране окончания аксона. Так как концентрация кальция снаружи клетки превышает концентрацию внутри клетки, кальций устремляется внутрь нервного окончания. В результате этого синаптичесие пузырьки сливаются с пресинаптической мембраной, а нейромедиатор путем экзоцитоза выводится в синаптическую щель. Молекула нейромедиатора диффундирует через синаптическую щель связываются со специфическими рецепторами в постсинаптической мембране. Это каналообразующие рецепторы, имеющие снабженные воротами каналы для натрия. Эти каналы открываются при связывании нейромедиатора с рецептором. Натрий устремляется внутрь клетки. Происходит деполяризация постсинаптической мембраны. Волна деполяризации распространяется по мембране нейрона. Молекулы нейромедиатора удаляются с помощью ферментов. В дальнейшем происходит обратный захват нейромедиатора или продуктов его разложения. Деполяризация (сдвиг потенциала в положительном направлении) постсинаптической мембраны характерна для возбуждающего синапса. Гиперполяризация (сдвиг потенциала в отрицательном направлении) - для тормозного синапса. 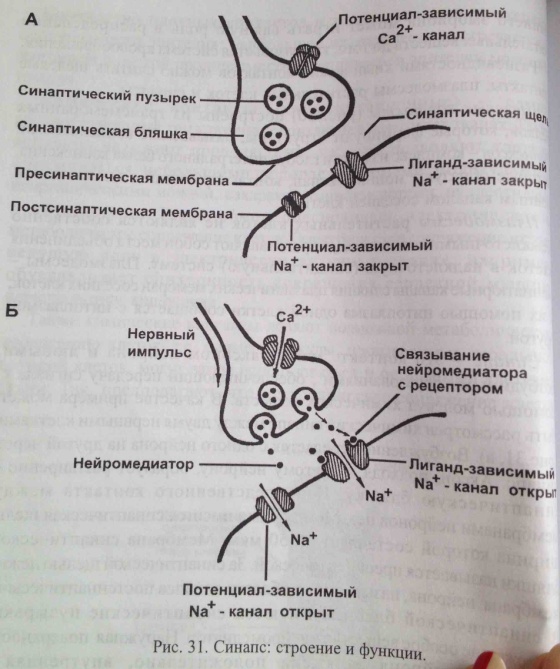 26. Синаптическая передача нервных импульсов. См. 25 вопрос. 27. Межклеточные соединения. Плотные или замыкающие контакты. Плотные контакты формируются за счет слипания глобул интегральных белков плазматических мембран соседних клеток. Белки укрепляются системой тонких фибрилл в цитоплазме, располагающихся параллельно поверхности клеток. Они обеспечивают одну из функций различных эпителиев-их работу с высокоизбирательной проницаемостью. Во-первых, они изолируют межклеточное вещество от внешней среды, настолько герметично соединяя клетки, что через образовавшийся клеточный слой не могут пройти даже малые молекулы. Во-вторых, они препятствуют перемещению белковых молекул в пределах липидного бислоя плазмалеммы. Это важно для поддержания направленности транспорта в клетке. Так, в клетках кишечного эпителия апикальная поверхность обращена в просвет кишки, белки, находящиеся здесь, обеспечивают избирательное поступление веществ из просвета кишки в клетку. Мембрана базальной стороны клеток содержит белки, откачивающие молекулы этих вществ из клетки в кровь. Наличие плотных контактов делает невозможным перемещение транспортных белков с апикальной стороны на базальную и наоборот, обеспечивая направленность транспорта. Количество плотных контактов зависит от степени проницаемости эпителия. 28. Метаболический аппарат клетки. Этапы внутриклеточного транспорта. Метаболизм (от греч. metabole — перемена, превращение), совокупность химических реакций, протекающих в живых клетках и обеспечивающих организм веществами и энергией для его жизнедеятельности, роста, размножения. Всключает ассимиляцию(анаболизм) и диссимиляцию(катаболизм) Анаболизм = Анаболический процесс - физиолого-биохимические процессы, направленные на усвоение клеткой пищевых веществ. В ходе анаболизма создается тело клетки. При этом химически чуждые соединения превращаются в соединения специфические для того или иного вида или особи. Метаболический аппарат клетки - совокупность ее структур, участвующих в метаболизме. Органоиды, участвующие в ассимиляции: (по каждому органоиду коротко - строение и функции) 1) шЭПС 2) глЭПС 3) аппарат Гольджи 4) рибосомы 5) хлоропласты (у растений) Диссимиляция существляется в: (по каждой структуре - коротко стороение и функции) 1) лизосомах (подготовительный этап) 2) в цитоплазме (гликолиз) 3) в митохондриях (аэробный этап) Митохондриальная ДНК образует собственный геном митохондрий, на который приходится около 1 % общего содержания ДНК в клетке. Кристы внутренней мембраны митохондрий большинства клеток имеют пластинчатую форму и лишь у некоторых клеток содержатся везикулярные или трубчатые кристы (в эндокринных клетках, продуцирующих стероидные гормоны). Функции: -Энергетические станции клетки (синтез АТФ на пластинчатых кристах). -Дыхательный центр клетки. -Участие в биосинтезе стероидов (на везикулярных кристах, с участием кислорода). -Участие в окислении жирных кислот. -Участие в синтезе нуклеиновых кислот 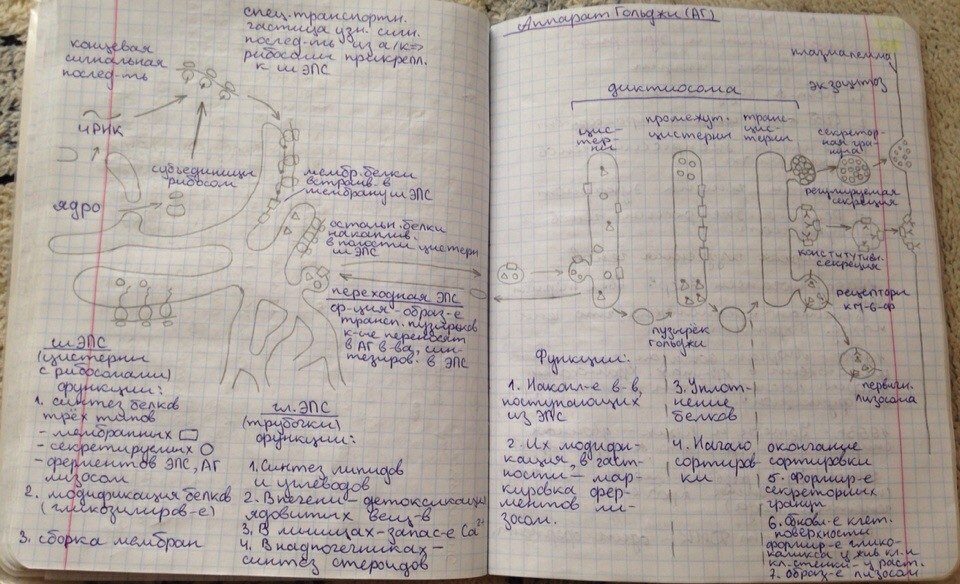 29. Современные представления о механизмах внутриклеточного транспорта. В клетках эукариот транспорт и локализация органелл, мембранных везикул осуществляется вдоль микротрубочек и актиновых микрофиламентов, образующих в них так называемый цитоскелет. Транспорт обеспечивают механохимических АТФазы, называемые моторными белками. Они, связанные с микротрубочками, относятся к двум большим семействам – кинезинам и динеинам, а с актиновыми микрофиламентами – к миозинам. Движение отдельных -результат действия нескольких моторных белков, и правильность транспорта обеспечивается тонкой регуляцией их активности. 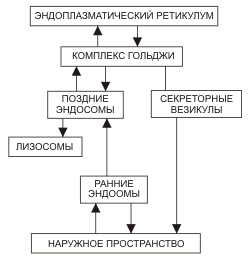 Везикулярный транспорт Из одной органеллы в другую перемещение происходит в везикуле или на ее поверхности в виде интегральных белков. Механизм внутриклеточного транспорта основан на взаимодействии т.с. с донорными и акцепторными компартментами. Выявлены белковые факторы узнавания пузырьком своей мишени (акцептор - компартмент) На каждом этапе осуществляется сортировка. Синтезируемые в цитоплазме на рибосомах белки должны попадать в разные компартменты клетки — ядро, митохондрии, ЭПР, АГ, лизосомы и др., а некоторые белки должны попасть во внеклеточную среду. Для попадания в определённый компартмент белок должен обладать специфической меткой. В большинстве случаев такой меткой является часть аминокислотной последовательности самого белка (лидерный пептид, или сигнальная последовательность белка). В некоторых случаях меткой служат посттрансляционно присоединённые к белку олигосахариды. Транспорт белков в ЭПР осуществляется по мере их синтеза, так как рибосомы, синтезирующие белки с сигнальной последовательностью для ЭПР, «садятся» на специальные транслокационные комплексы на мембране ЭПР. Из ЭПР в АГ, а оттуда в лизосомы, на внешнюю мембрану или во внеклеточную среду белки попадают путём везикулярного транспорта. В ядро белки, обладающие сигнальной последовательностью для ядра, попадают через ядерные поры. В митохондрии и хлоропласты белки, обладающие соответствующими сигнальными последовательностями, попадают через специфические белковые поры-транслокаторы при участии шаперонов. Пузырьки из Эпр-цистерны аппарата гольджи,сначала проходит через цис цистерну(, синтез),в промежут цистерну(сортировка),в транс(окончат сортировка и вывод из кл). Донорый компартмент – органелла от которой отрывается мембрана в составе везикулы, акцепторный компартмент – принимает везикулу. конститутивная секреция – происходит постоянно и не зависит от внешних сигналов. регулируемая секреция – под ПМ происходит накопление пузырьков, которые сливаются с ПМ при наличии внешних сигналов – гормоны, нервы – и повышении конц. Ca2+ до 1мкм ретроградный транспорт – возвращение рецепторных белков и липидов из АГ в Эр - восполнение мембраны ЭР. 30. Этапы энергетического обмена в клетке. Анаэробный этап. Брожение. Первый этап — подготовительный. Пища поступает в организм животных и человека в виде сложных высокомолекулярных соединений. Прежде чем поступить в клетки и ткани, эти вещества должны разрушиться до низкомолекулярных, более доступных для клеточного усвоения веществ. На первом этапе происходит гидролитическое расщепление органических веществ, идущее при участии воды. Оно протекает под действием ферментов в пищеварительном тракте многоклеточных животных, в пищеварительных вакуолях одноклеточных, а на клеточном уровне — в лизосомах. Второй этап осуществляется на клеточном уровне при отсутствии кислорода. Он протекает в цитоплазме клетки. Рассмотрим расщепление глюкозы, как одного из ключевых веществ обмена в клетке. Все остальные органические вещества (жирные кислоты, глицерин, аминокислоты) на разных этапах втягиваются в процессы ее превращения. Бескислородное расщепление глюкозы называется гликолизом. Третий этап — биологическое окисление, или дыхание. Этот этап протекает только в присутствии кислорода и иначе называется кислородным.Он протекает в митохондриях. Начальные этапы окисления углеводов происходят в цитозоле и не требуют участия кислорода, поэтому эта стадия процесса называется анаэробным окислением, или гликолизом. Главным субстратом окисления при анаэробном получении энергии служат гексозы и в первую очередь глюкоза. Процесс гликолиза включает 9 последовательных ферментативных реакций, в результате которых молекула глюкозы превращается в две молекулы пирувата. В ходе некоторых из этих реакций происходит восстановление НАД+ до НАДН и перенос неорганического фосфата на АДФ с образованием высокоэнергетической связи АТФ. В глюкозе количество потенциальной энергии, заключенной в связях между атомами С, Н и О, составляет около 680 ккал на 1 моль. Эта энергия освобождается при полном окислении глюкозы: С6Н12О6+6О2=6Н2О+6СО2+680ккал. В процессе гликолиза происходит неполное окисление субстрата. В результате него глюкоза распадается до триоз, при этом тратятся 2 молекулы АТФ и синтезируются 4 молекулы АТФ, так что в конечном результате клетка получает всего 2 молекулы АТФ. В энергетическом отношении этот процесс малоэффективен, поэтому из 680 ккал, заключающихся в связях 1 моля глюкозы, освобождается менее 10% энергии. Несмотря на низкий энергетический выход, анаэробное окисление, гликолиз, широко используется в живой природе. Эритроциты млекопитающих, например, получают всю необходимую им энергию за счет гликолиза, так как у них нет митохондрий. Конечные продукты гликолиза - триозы, в первую очередь пировиноградная кислота, все еще несут большое количество химической энергии и вовлекаются в дальнейшее окисление, происходящее в самих митохондриях. 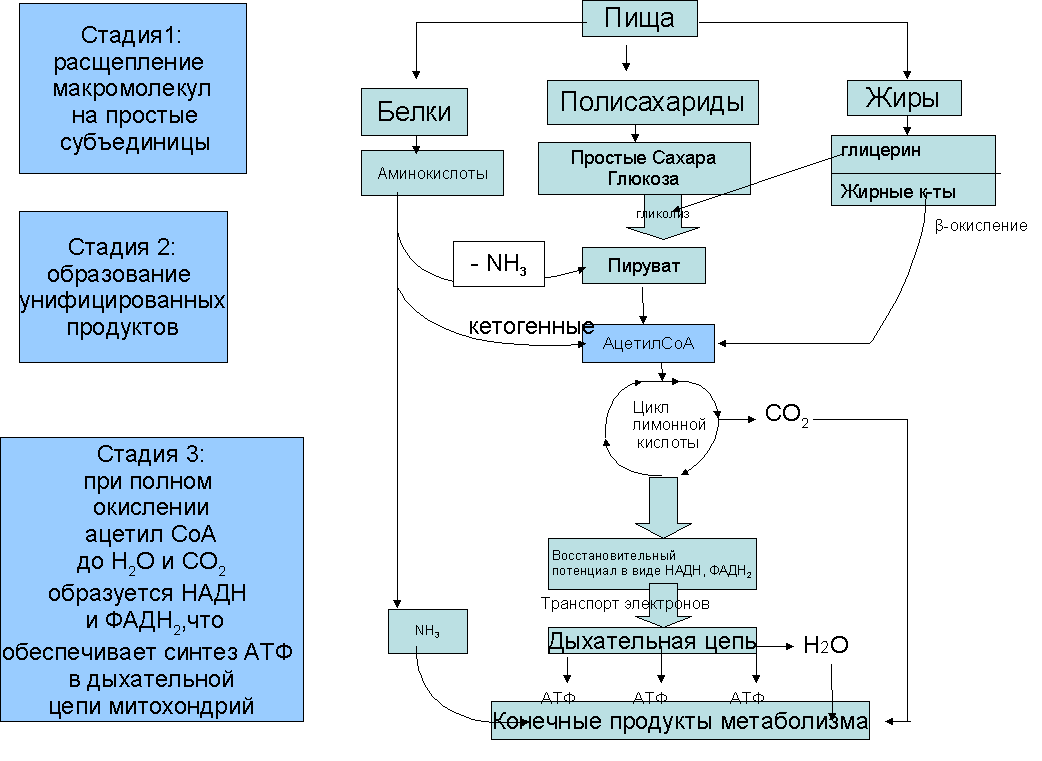 31. Этапы энергетического обмена в клетке. Аэробный этап. Про этапы см. 30 вопрос. Окислительные процессы в митохондриях связаны с окислительным циклом трикарбоновых кислот и с дыхательной цепью переноса электронов. Они начинаются после образования в матриксе достаточного количества ацетил-КоА из пирувата и жирных кислот. Пируват, образовавшийся в результате гликолиза, перемещается из цитозоля в матрикс митохондрии, где теряет молекулу СО и, окисляясь до ацетата, соединяется с коферментом А. Жирные кислоты в митохондриальном матриксе окисляются, молекула жирной кислоты последовательно укорачивается на два углеродных атома, и образуется ацетил-КоА. Ферменты, катализирующие эти реакции, как и большинство ферментов цикла трикарбоновых кислот, находятся в матриксе митохондрий. Главная функция цикла лимонной кислоты-окисление ацетата, который включается в цикл в форме молекул ацетил-КоА. Цикл начинается взаимодействия ацетил-КоА с оксалоацетатом, что приводит к образованию цитрата. Далее в ходе семи последовательных ферментативных реакций два атома углерода удаляются в виде углекислого газа, и в конце концов регенерируется оксалоацетат. Каждый оборот цикла дает две молекулы углекислого газа, который диффундирует из митохондрий и покидает клетку. Энергия, высвобождающаяся при окислении химических связей лимонной кислоты, потребляется в цикле трикарбоновых кислот несколькими способами. Во-первых, в одной из реакций цикла синтезируется одна молекула АТФ. Во-вторых, в ходе реакций цикла лимонной кислоты происходит извлечение высокоэнергетических электронов, которые используются для восстановления молекул-переносчиков водорода НАД (никотинамидадениндинуклеотида) и ФАД (флавинадениндинуклеотида). В каждом обороте цикла три молекулы НАД превращаются в НАДН и одна молекула ФАД-в ФАДН. Образование НАДН происходит не только в ходе реакций цикла лимонной кислоты, но и на более ранних этапах окисления веществ: в процессе гликолиза и при образовании ацетил-КоА из пирувата. Энергия, переносимая молекулами НАДН и ФАДН далее используется в реакциях окислительного фосфорилирования для синтеза АТФ. Окислительное фосфорилирование связано с переносом электронов от НАДН и ФАДН по дыхательной цепи к молекулярному кислороду и использованием энергии для синтеза АТФ. Эти процессы протекают на внутренней мембране митохондрий, в которой находятся белки-переносчики дыхательной цепи и комплексы АТФ-синтетазы. Дыхательная цепь внутренней мембраны митохондрий содержит три главных ферментных комплекса, участвующих в переносе электронов с НАДН на кислород. Первый, НАДН-дегидрогеназный, комплекс принимает электроны от HAДH и переносит их во второй комплекс-комплекс цитохромов b-с, который переносит их на цитохромоксидазный комплекс, передающий электроны на кислород, в результате чего образуется вода. На этом процесс окисления заканчивается (конечными продуктами окисления исходного субстрата являются углекислый газ и вода). Перенос электронов по дыхательной цепи происходит с участием убихинона и цитохрома с-мобильных переносчиков, передвигающихся в мембране от одного комплекса к другому и обратно. Переносчики электронов расположены и дыхательной цепи таким образом, что каждый последующий комплекс обладает большим сродством к электронам, чем предыдущий. Электроны перемещаются от одного комплекса к другому, пока не перейдут на кислород, имеющий наибольшее сродство к электрону. Перенос электронов сопровождается их переходом на более низкие энергетические уровни и выделением энергии. Элементы дыхательной цепи последовательно окисляются и восстанавливаются, в результате чего энергия высвобождается небольшими порциями и используется для перекачивания протонов из матрикса митохондрии через внутреннюю митохондриальную мембрану в межмембранное пространство и далее за пределы митохондрии. Перенос протонов, осуществляемый компонентами дыхательной цепи, приводит к возникновению электрохимического протонного градиента, который складывается из мембранного потенциала и градиента pH. Энергия электрохимического протонного градиента используется для синтеза АТФ. Преобразование энергии окислительных реакций в энергию АТФ происходит на внутренней митохондриальной мембране в результате хемиосмоса. При достижении определенной разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс и при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. АТФ-синтетаза состоит из двух основных частей: мембранного компонента F0, осуществляющего транспорт протонов через мембрану, и сопрягающего фактора F1, осуществляющего синтез АТФ. Окислительные процессы в митохондриях и фосфорилирование АДФ можно разобщить, сняв разность потенциалов на митохондриальной мембране, сделав в ней диффузионные каналы, а также механическим нарушением, либо с помощью различных химических соединений. Перенос электронов и окисление субстрата будут продолжаться, но синтеза АТФ не будет. Разобщение метаболических процессов окисления и фосфорилирования приводит к образованию значительного количества тепла вместо накопления энергии в форме макроэргических соединений. Суммарное уравнение: С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 38АТФ + Q |