1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
43. Опишите путь макромолекулы от момента проникновения в клетку до её усвоения. 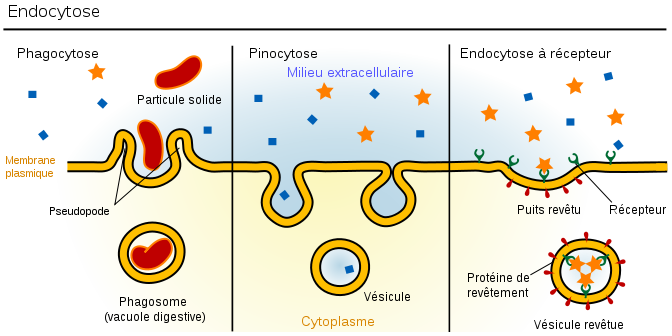 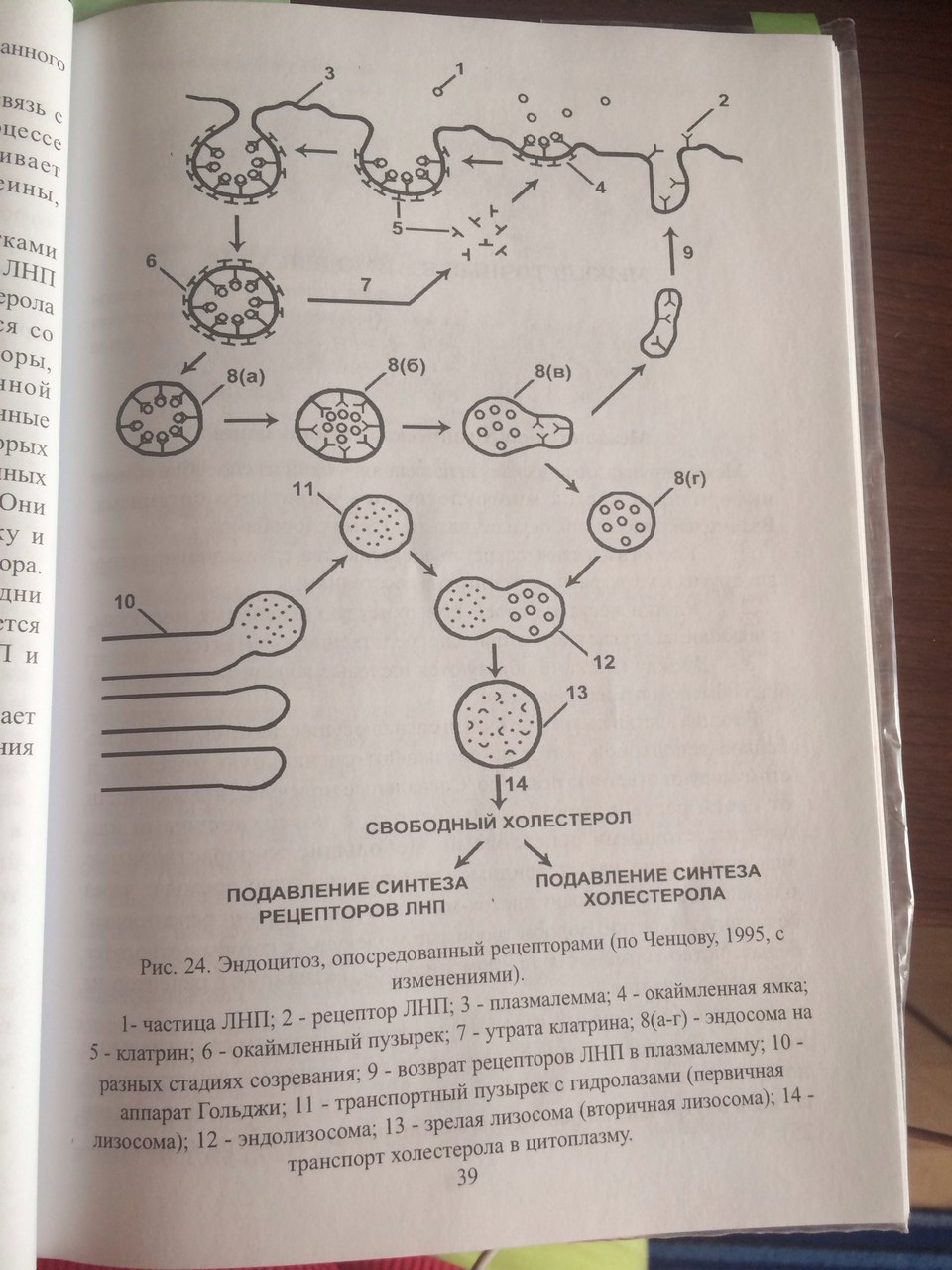 Эндоцитоз - это образование пузырьков путем выпячивания плазматической мембраны при поглощении твердых частиц-макромолекул (фагоцитоз) или растворенных веществ (пиноцитоз). Возникающие при этом гладкие или окаймленные эндоцитозные пузырьки называют также фагосомами или пиносомами. Путем эндоцитоза осуществляются: -питание (яйцеклетки поглощают таким способом желточные белки, фагосомами являются пищеварительные вакуоли простейших); -защитные и иммунные реакции (лейкоциты поглощают чужеродные частицы и иммуноглобулины); -транспорт (почечные канальцы всасывают белки из первичной мочи). -поглощение ЛНП(липопротеинов низк плотн) (Поглощение животными клетками холестерола из внеклеточной среды осуществляется за счет эндоцитоза, опосредуемого рецепторами. Основная часть холестерола переносится кровью в виде ЛПНП . Молекулы ЛНП связываются со специальными рецепторами в плазмолемме клетки рецепторы, соединённые с лигандом(ЛНП) перемещаются по клеточной поверхности и накапливаются в окаймлённых ямках(их субмембранный аппарат содержит клатрин) из ямок обр-ся путём впячивания эндоцитозные пузырькипоступают в цитоплазму, теряют клатриновую оболочкусливаются с эндосомой, где лиганд отделяется от рецептораот мембраны эндосомы отделяются 2 вида пузырьков:одни содержат ЛНП и сливаются с лизосомой, где из ЛНП выщепляется холестерол, другие содержат свободные рецепторы, которые возвращаются в мембрану.) Избирательный эндоцитоз определенных веществ (желточных белков, иммуноглобулинов и т. п.) происходит при контакте этих веществ с субстрат-специфическими рецепторными участками на плазматической мембране. Субстраты, попадающие в клетку путем эндоцитоза, расщепляются ("перевариваются"), накапливаются (например, желточные белки) или снова выводятся с противоположной стороны клетки путем экзоцитоза. 44. Роль ЭР и АГ в регенерации и обновлении поверхностного аппарата клетки(ПАК). На рибосомах гранулярного ЭПР происходит синтез белков, которые в зависимости от их конечного назначения могут быть разделены на три группы: 1.секреторные белки. 2.внутриклеточные белки(гидролазы) 3.мембранные белки. В регенерации плазмалеммы принимают участие мембранные белки, которые после синтеза остаются в составе мембраны ЭПР, «отпочковываясь» вместе с частью мембраны эти белки поступают в АГ, где происходит присоединение углеводных цепочек. Пройдя после цис-полюса через промежуточный полюс к транс- полюсу мембанные белки отправляются на «ремонт» мембраны. 45. Пероксисомы. Образование, строение и функции. Мембранные пузырьки с однородным или гранулярным матриксом. В центре-сердцевина (нуклеоид) из кристаллических структур, образованных фибриллами или трубочками. Во фракции пероксисом обнаруживается до 15 ферментов, связанных с метаболизмом перекиси водорода (Н2О2). Это окислительные ферменты, при работе которых образуется перекись водорода, и каталаза, приводящая к утилизации токсичной для клеток Н2О2. Таким образом, пероксисомы защищают клетку от действия перекиси водорода. Пероксисомы обнаружены во всех эукариотических клетках. Они являются главным центром утилизации кислорода (наряду с митохондрией). Кислород используется ими для окислительных реакций, не сопровождающихся накоплением энергии в виде АТФ. Образующаяся перекись водорода используется для окисления субстратов, а ее излишки разрушаются с образованием воды. Число пероксисом варьирует в клетках разных типов. Особенно важное значение они имеют в растительных клетках. В прорастающих семенах имеется разновидность пероксисом-глиоксисомы. Они обеспечивают превращение жирных кислот запасенных липидов в сахара в ходе реакций глиоксилатного цикла. Другой тип пероксисом в клетках листьев катализирует окисление побочных продуктов фотосинтеза. Пероксисомы способны к саморепродукции. Новые образуются в результате роста и деления предшествующих. Белки поступают в пероксисому из цитозоля. Липиды переносятся в их мембраны от места синтеза через цитозоль. Доставка липидов обеспечивается специальными транспортными белками-переносчиками. 46. Организация цитоскелета. Система микрофиламентов. Микрофиламенты-тонкие белковые нити толщиной около 8 нм, построенные из белка актина. Актин присустствует в двух состояниях: в виде отдельных глобулярных субъединиц или в виде филаментов, образованных в результате полимеризации глобулярного актина. Микрофиламенты полярны: они имеют плюс-конец, который растет за счет полимеризации актина, и минус-конец, где происходит деполимеризация. Микрофиламенты пронизывают всю цитоплазму клетки, образуя сложную сеть, которую поддерживают специальные сшивающие белки. Под плазматической мембраной эта сеть наиболее густая и образует клеточный кортекс, который придает прочность поверхностному слою клетки и позволяет ей изменять свою форму и двигаться. Например, когда фагоцитирующий лейкоцит вступает в контакт с бактерией, сеть актиновых филаментов в этом участке кортекса распадается, цитоплазма превращается из геля в золь. Золь позволяет поверхностному слою цитоплазмы окружить и поглотить бактериальную клетку. Такое временное разжижение цитолазмы ссвязано с действием белка гельзолина. Течение цитоплазмы и движение немышечных клеток может происходить при взаимодействии немышечного миозина с актиновыми филаментами. В мышечных клетках присутствует мышечный миозин, который образует крупные фибриллы (в отличие от немышечного). 47. Сократительные структуры в клетке. Механизм мышечного сокращения. Длинные тонкие мышечные волокна, из котрых построена скелетная мышца, -многоядерные гигантские клетки, образующиеся в ходе онтогенеза при слиянии множества отдельных клеток. Мышечные волокна содержат миофибриллы. Пучки миофибрилл имеют поперечную исчерченность. Темные участки (А-диски)-анизотропные, светлые (I-диски)-изотропные. По середине светлого I-диска – Z-диск. Единицей строения миофибриллы- саркомер (участок между двумя Z-дисками). В сотав саркомера входят тонкие и толстые филаменты (нити). Толстые построены из блка миозина, располагаются в А-дисках. Молекула миозина имеет хвост и головку, способную расщеплять АТФ и взаимодействовать с актином. В толстых нитях молекулы миозина головками обращены в противоположные стороны, хвостами-друг к другу. Тонкие филаменты присутствуют в I-дисках, одним концом закреплены на Z-дисках. Сформированы актином. Содержат тропонин и тропомиозин. Сокращение миофибриллы происходит за счет уменьшения длины саркомеров. В основе-скольжение тонких и толстых нитей относительно друг друга. При этом миозиновые филаменты входят в пространства между актиновыми, приближая друг к другу Z-диски. В покоящейся мышце длинные филаменты тропомиозина закрывают центры связывания с миозином на молекулах актина. Сокращение скелетной мышцы начинается после поступления нервного импульса. Из саркоплазматического ретикулума выходят ионы кальция, которые связываются с тропонином тонких филаментов. Тропонин меняет свою конформацию и смещает тропомиозин, открывая на актиновых филаментах центры связывания с миозином. Это делает возможным взаимодействие головок миозина с актином. Головка миозина с присоединенным к ней АДФ и Ф имеет конформацию, позволяющую связываться с актином. Связывание с актином вызывает изменеие конформации головки: она наклоняется и тянет актиновый филамент к центру саркомера, освобождаясь от АДФ и Ф. Далее присоединение АТФ приводит к отделению ее от актина. Головка миозина гидролизует АТФ до АДФ и Ф и принимает исходную конформацию. 48. Организация цитоскелета. Система промежуточных филаментов. Промежуточные филаменты-прочные белковые волокна, пронизывающие цитоплазму. Их структура напоминает переплетенные канаты, толщина 8-10 нм. В большинестве животных клеток они формируют характерную «корзинку» вокруг ядра, откуда тянутся к периферии клетки. Особенно их много там, где клетки подвергаются механическим нагрузкам. В клетках разных тканей они построены из разных белков. Несколько видов кератинов формируют промежуточные филаменты в эпителиальных клетках и производных эпидермиса. Виментин распространен в клетках мезенхимного происхождения. Десмин содержится в мышечных клетках. 49. Организация цитоскелета. Система микротрубочек. Производные микротрубочек. Микротрубочки-полые цилиндры, образованные молекулами белка тубулина. Каждая молекула тубулина-это димер, образованный двумя субъединицами: альфа- и бета-тубулином. При формировании микротрубочек молекулы тубулина соединяются друг с другом в протофиламенты-нитевидные комплексы, в которых альфа-тубулин одного димера контактирует с бета-тубулином следующего. Микротрубочка модеожит 13 таких протофиламентов., расположенных параллельно и примыкающих друг к другу. Микротрубочки являются полярными структурами: наличие быстро растущего плюс-конца и медленно растущего минус-конца. Удлинение происходит путем добавления молекул тубулина. При недостаточной концентрации тубулина микротрубочки могут разбираться с обоих концов. Рост микротрубочек регулируют клеточный центр и базальные тельца. Они могут либо не образовывать структур, либо формировать временные образования (например, веретено деления) или могут входить в состав постоянных клеточных структур. Цитоплазматические микротрубочки существуют в виде отдельных нитей. Они образуют систему волокон, по которой могут перемещаться органоиды. Они влияют на полярность клетки, регулируют ее форму и движение и определяют ориентацию плоскости клеточного деления. Клеточный центр-центр образования микротрубочек. Расположен около ядра и имеет пару центриолей, расположенных под прямым углом друг к другу. Центриоли окружены зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросфера). Центриоль представляет собой цилиндр, стенка которого образована девятью триплетами микротрубочек. Соседние триплеты соединены между собой белковыми сшивками, поддерживающими цилиндрическую структуру. Клеточный центра характерен для клеток животных. В делящихся клетках центриоли участвуют в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках клеточный центр часто определяет полярность клеток. 50. Специализированные структуры плазматической мембраны (микроворсинки, реснички и жгутики).  Внутри реснички(жгутика) расположена аксонема-сложная структура, состоящая в основном из микротрубочек. В аксонеме девять сдвоенных микротрубочек расположены по окружности, в центре которой-две одинаковые микротрубочки. Такая структура «(9х9+2» характерна для ресничек и жгутиков почти всех эукариотических организмов. Аксонема стабилизируется вспомогательными белками. Например, белок нексин образует центральный опорный цилиндр вокруг пары центральных микротрубочек и радиальные спицы, связывающие центральный цилиндр с периферическими дублетами микротрубочек. Он также образует между соседними дублетами поперечные связи, которые стягивают аксонему по окружности. От каждого дублета микротрубочек отходят короткие боковые выступы, образованные динеином-динеиновые ручки. Они образуют временные контакты со стенкой микротрубочки соседнегодублета и играют важную роль в движении ресничек и жгутиков. Взаимодействие тубулина и динеина вызывает скольжение дублетов относительно друг друга и изгибание ресничек. В основании реснички-центриоль (базальное тельце). Две из трех микротрубочек непосредственно переходят в микротрубочки периферического дублета аксонемы. Базальное тельце погружено в цитоплазму, специальные отростки соединяют его с другими компонентами цитоскелета. Оно служит опорой ресничке и организует рост микротрубочек аксонемы при образовании или регенерации реснички. 51. Образование и роль рибосом в клетке. В клетках эукариот существует 2 разновидности рибосом: рибосомы цитоплазмы (80S) и рибосомы, находящиеся в митохондриях и пластидах (50-80S) Рибосомы состоят из двух субъединиц большой и малой. Малая субъединица рибосомы удерживает мРНК и тРНК, а большая катализирует образование пептидной связи. В состав субъединиц входят рРНК и белки (преимущественно глобулярные). Большая субъединица рибосомы состоит из 3 молекул рРНК и 50 белков, а малая из 1 молекулы рРнк и 30 белков. Субъединицы рибосом собираются в ядрышке и через ядерные поры выходят в цитоплазму, где находятся в диссоциированном состоянии. 2 субъединицы образуют комплекс- рибосому только при осуществлении синтеза полипептида на иРНК(процесс трансляции). Если к иРНК прикрепляется множество рибосом, то такой комплекс называется полисомой. 52. Морфология ядерных структур. В ядерном аппарате эукариотической клетки выделяют следущие части: поверхностный аппарат ядра, ядерный матрикс, кариоплазму, хроматин и ядрышко. Поверхностный аппарат: ядерная оболочку с поровым комплексом и ламина. Ядерная оболочка образована наружной и внутренней ядерными мембранами, между которыми перинуклеарное пространство. Наружная переходит во внутреннюю в области ядерных пор. Наружная мембрана переходит в мембраны ЭПР, а перинуклеарное пространство таким образом оказывается связанным с полостью каналов и цистерн ЭПР. Поровый комплекс представляет собой 2 кольца из 8 белковых глобул, расположенных по краю порового отверстия- в области слияния наружной и внутренней мембран. В центре поры может быть видна центральная гранула. Поровый комплекс представляет собой надмолекулярную структуру . Они обладают собственными рецепторами, аппаратом, регулирующим направление, способ и интенсивность транспорта через пору. Ламина - плотная пластинка, примыкающая к внутренней мембране ядерной оболочки. Представляет собой густую сеть белковых фибрилл. Ламина способствует поддержанию формы ядра, выполняет функцию упорядочения расположения интерфазных хромосом, связана с цитоселетом и играет важную роль в поддержании порового комплекса. Ядерный матрикс-«Скелет» ядра. Представляет собой комплекс фибриллярных белков, который обеспечивает структурную организацию всех компонентов ядра и участвует в регуляции процессов репликации, транскрипции, созревании продуктов транскрипции, выведении их в цитоплазму. С ядерным матриксом связан актин, который, видимо, участвует в ругеляции транскрипции. Кариоплазма(ядерный сок) создает специфическое для ядерных структур микроокружении, обеспечивая возможность их нормального функционирования. Ядрышкопредставляет собой структуру в которой происходит образование рибосомальных единиц. Здесь находятся участки ДНК, содержащие многочисленные одинаковые гены рРНК. В метафазных хромосомах эти участки (ядрышковые организаторы) локализованы в области вторичной перетяжки. У человека они находятся в 13,14,15,21,22 хромосомах. Кроме того, гены рРНК находятся также в 1 паре хромосом. Различают фибриллярный и гранулярный компоненты ядрышка. Фибриллярная зона содержит ДНК, рРНК, а гранулярная часть-зона со зрелыми субъединицами рибосом. Хроматин- (ДНП-дезоксирибонуклеопротеид) состоит из ДНК, РНК и белков (гистонов-основных, негистонов-кислых) хроматин составляет основу хромосом. 53. Роль ядерных структур в жизнедеятельности клетки. В ядерном аппарате эукариотической клетки выделяют следующие части: поверхностный аппарат ядра, ядерный матрикс, кариоплазму, хроматин и ядрышко. Функции поверхностного аппарата: 1.ядерная оболочка отграничевает ядерное пространство от цитоплазмы, что позволяет разграничить во времени и пространстве этапы транскрипции и трансляции биосинтеза белка. В результате стал возможен процессинг- этап преобразования первичного транскрипта, приводящий к образованию зрелой РНК. 2.ядерная оболочка защищает ДНК от механических воздействий цитоскелета цитоплазмы 3.через поровый комплекс осуществляется двусторонний транспорт. Функции ядерного матрикса: обеспечивает структурную организацию всех компонентов ядра и участвует в регуляции процессов репликации, транскрипции, созревании продуктов транскрипции, выведении их в цитоплазму. Функции ядрышка: представляет собой структуру в которой происходит образование рибосомальных единиц. Функция кариоплазмы: создание специфического для ядерных структур микроокружении, обеспечивая возможность их нормального функционирования. Ядро осуществляет две группы общих функций: одна из них — хранение генетической информации, другая — ее реализация, обеспечение синтеза белка. В первую группу входят процессы, обусловливающие поддержание наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК, что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее в ядре происходят воспроизведение, или редупликация, и разъединение молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном и количественном смысле объемы генетической информации. В ядре эукариот происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток. Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственного аппарата белкового синтеза. Это не только синтез (транскрипция) на молекулах ДНК разных информационных РНК, но также транскрипция всех видов трансферных РНК и рибосомных РНК. В ядрах эукариотических клеток происходит «созревание» (процессинг, сплайсинг) первичных транскриптов. В ядре эукариот образуются также субъединицы рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро. Таким образом, ядро представляет собой не только вместилище генетического материала, но и структуру, где этот материал воспроизводится и функционирует. Поэтому выпадение или нарушение любой из перечисленных выше функций гибельно для клетки в целом. |