1. о сущности живого. Нуклеопротеидные комплексы. Эволюция представлений о химической сущности жизни. Ф. Энгельс Жизнь способ существования белковых тел
 Скачать 17.11 Mb. Скачать 17.11 Mb.
|

|
32. Гипотеза эволюционного происхождения митохондрий, хлоропластов, ядра, ЭПР. Возникновение клеточной организации в ходе эволюции. (нет) Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 годуАндреас Шимпер, показавший их саморепликацию внутри клетки. А. С. Фаминцин и О. В. Баранецкий-о двойственной природе лишайников - комплекса гриба и водоросли. К. С. Мережковский в 1905 году предложил само название «симбиогенез», впервые детально сформулировал теорию и даже создал на её основе новую систему органического мира. Фаминцин в 1907 году, опираясь на работы Шимпера, также пришёл к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников. В 1920-е Б. М. Козо-Полянский- симбионтами являются и митохондрии. Митохндрии — это потомки аэробных бактерий (прокариот),поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов. Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли. Доказательства Митохондрии и пластиды: • имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий. • размножаются бинарным делением (причём делятся иногда независимо от деления клетки) • генетический материал — кольцевая ДНК, не связанная с гистонами (ДНКмитохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот) • имеют свой аппарат синтеза белка — рибосомы и др. Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза (англ. viral eukaryogenesis). В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра. 33. Митохондрии. Организация потока энергии в клетке. 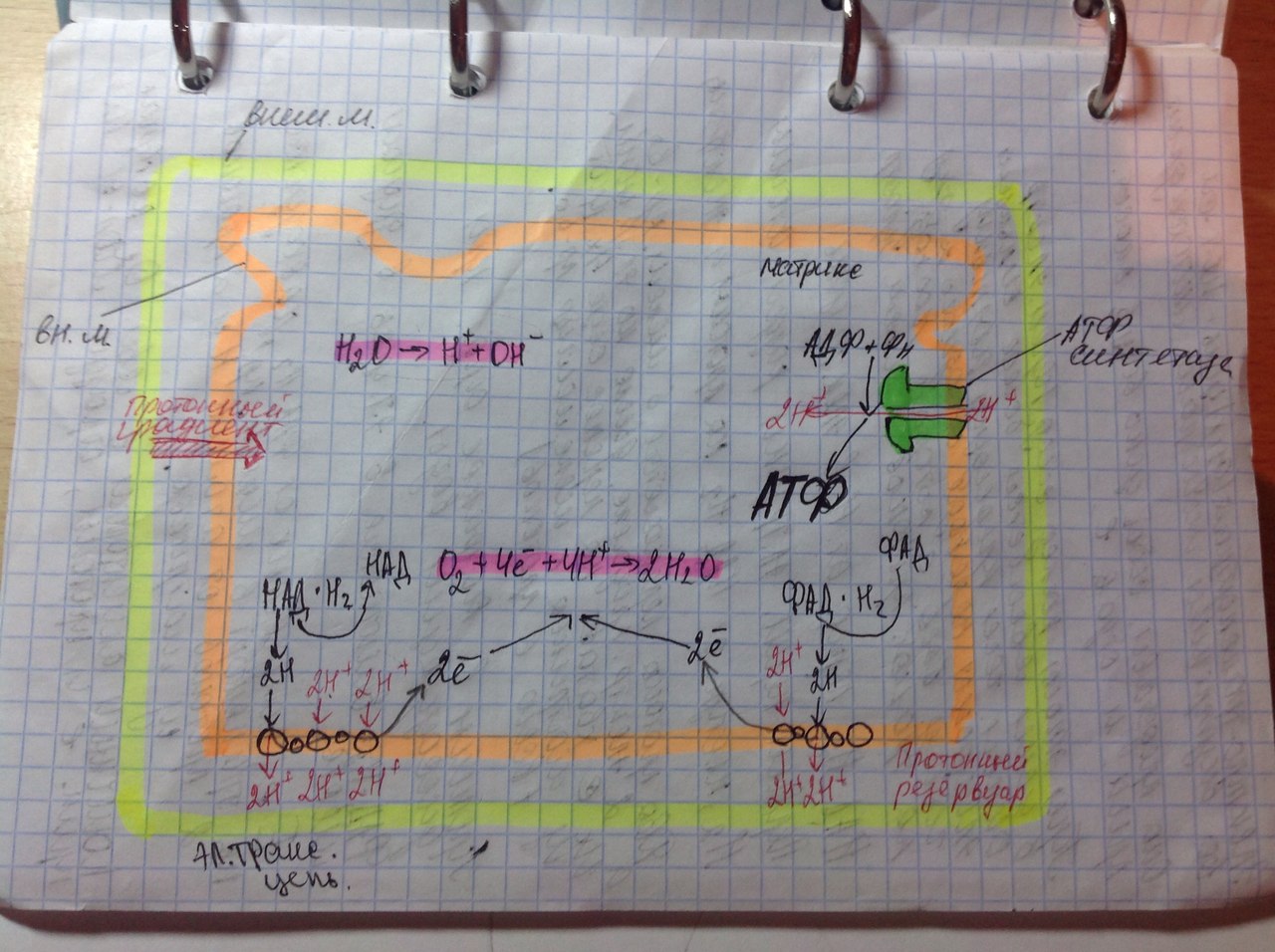 Митохондрии -двухмембранные органеллы эукариотических клеток, обладающие собственной ДНК(испыт повреждения, тк нах в матриксе где много биоокислителей-митохондриальн.болезни) и выполняющие функцию синтеза АTФ. Митохондрии расположены около мест высокого потребления АТФ(в скелетных мышцах - вблизи миофибрилл. В сперматозоидах - футляр вокруг оси жгутика. В аксонах нервных кл-около синапсов, где происходит процесс передачи нервного импульса) Поток энергии обеспечивается механизмами энергоснабжения —брожением, фото- или хемосинтезом, дыханием. Центральная роль в биоэнергетике клеток животных принадлежит дыхательному обмену. Он включает реакции расщепления глюкозы, жирных кислот, аминокислот, а также использование выделяемой энергии для образования «топлива» в виде аденозинтрифосфата (АТФ). Особое место в дыхательном обмене принадлежит митохондриям, выполняющим функцию окислительного фосфорилирования. Недоокисленные продукты гликолиза (пируват) поступают в матрикс митохондрий, где в условиях полного окисления, сопряженного с фосфорилированием АДФ до АТФ, отдают для нужд клетки оставшуюся в их химических связях энергию. Взаимосвязь внутриклеточного обмена белков, жиров и углеводов обеспечивает цикл Кребса. Ацетил-Коа взаимод с оксалоацетатом=цитрат. Далее в ходе 7-ми последовательных реакц 2 атома «С» удаляются в идее СО2, в конце концов регенерируется оксалоацетат. Высвобаждаются: 1АТФ и 1ФАДН2 с 3НАДН2, кот используются в окислительном фосфорилировании для синтеза АТФ путем перекачки протонов и электоронов. Окислительное фосфорилирование-внутр мембр митохондрий, где нах дых цепь(НАДН-дегидрогеназный компл, компл цитохромов b-c1,цитохромоксидазный компл) и комплекс АТФ-ситетазы. Электроны перемещ от одного компл к другому, достигая кислорода, имеющ наибольшее сродство к электрону. Переход на более низкие уровни сопровождается выделением энергии, использ для перекачки протонов в межмембранное пространство. Так возникает протонный градиент(мембр потенциал+градиент рН). При достижении определ разности потенциалов, АТФ-синтетаза начинает транспортировать протоны в матрикс, превращая АДФ и неорганич фосфат в энергию АТФ. ИТОГ: 34+2+2=38 АТФ Суть цикла: Запасение энергии в виде высокоэнергетическоемких электронов, входящих в соста атомов водорода, связ с НАД и ФАД, в электрон транспортную сеть. Суммарное уравнение может быть записано в следующем виде: Весь водород из молекулы глюкозы оказывается в конечном счете у переносчиков (НАД и ФАД). Весь углерод теряется в виде С02. 34. Образование АТФ в митохондриях. См. 31 вопрос 35. Образование АТФ в клетке. Хемиосмотическая гипотеза Митчела. См. 31 вопрос Хемиосмотическая теория: Для образования АТФ ферментная система АТФ-синтетаза использует разницу электрохимических потенциалов и концентрации ионов водорода на разных сторонах мембраны, перераспределяя поток водорода: с внешней мембраны переносит ионы водорода на внутреннюю. Во время перенесения электронов от НАДН к кислороду выделяется энергия, необходимая для синтеза трех молекул АТФ. 36. Гладкий ретикулум. Строение и функции. Гладкий ЭПР состоит из мембранных трубочек, канальцев, вакуолей. На его мембранах нет рибосом. Степень развития гладкого ЭПР неодинакова как в разных клетках, так и внутри одной клетки. Он весьма лабилен и способен к существенным перестройкам при изменяющихся условиях. Одной из важнейших функций гладкого ЭПР является синтез липидов (в том числе мембранных). Поэтому он преобладает в клетках, специализирующихся на метаболизме липидов. Деятельность гладкого ЭПР также связывают с метаболизмом некоторых внутриклеточных полисахаридов, в частности с синтезом гликогена. В некоторых клетках гладкий ЭПР специализируется на выполнении особых функций. Например, в клетках коры надпочечников в ЭПР синтезируются стероидные гормоны. Здесь располагаются ферменты, обеспечивающие синтез холестерола и его преобразование в стероидные гормоны. В гепатоцитах позвоночных гладкий ЭПР специализирован на детоксикации токсических веществ. Гладкий ЭПР при этом сильно разрастается. После удаления токсических веществ избыток сети уничтожается аутофагическим путем. В поперечно-полосатых мышцах гладкий ЭПР выполняет функцию депонирования ионов кальция. 37. Строение и функции гранулярного ретикулума. Гранулярный ЭПР представлен системой плоских цистерн. На их мембране со стороны цитозоля расположены рибосомы, объединенные в полисомы. На рибосомах гранулярного ЭПР происходит синтез белков, которые в зависимости от их конечного назначения могут быть разделены на три группы: 1.белки, предназначенные для секреции 2.белки внутренней фазы ЭПР, аппарата Гольджи, лизосом 3.мембранные белки, предназначенные для ЭПР, аппарата Гольджи, лизосом, ядерной оболочки и плазмалеммы. В ЭПР происходят начальные этапы сортировки синтезированных белков. Растворимые белки первых двух групп целиком поступают в цистерны ЭПР, который обеспечивает их обособления от цитозоля. Мембранные белки после синтеза остаются в составе мембраны ЭПР. Сборка любого белка начинается на свободных рибосомах в цитозоле. В ЭПР поступают только те пептиды, у которых первым синтезируется специфический гидрофобный сигнальный пептид. Особая частица, распознающая сигнал (SRP) связывается с сигнальным пептидом, временно блокирует синтез белка и затем направляет рибосому к мембране эндоплазматического ретикулума, где SRP присоединяется к своему рецептору. Доставленная таким образом к ЭПР рибосома прикрепляется своей большой субъединицей к специальному белку-рецептору, который участвует в образовании канала. Частица, выполнившая свою задачу, покидает рибосому. Прекращается ее блокирующее действие, и синтез белка возобновляется. Растущая белковая цепь поступает в ЭПР через канал в мембране. Пока белок в виде петли перетаскивается в полость ЭПР, его гидрофобный сигнальный пептид остается погруженным в мембрану. При синтезе растворимых белков сигнальный пептид отрезается, и белок высвобождается в полость ЭПР. Трансмембранные белки остаются заякоренными в билипидном слое с помощью неотрезанного сигнального пептида или за счет другого гидрофобного участка-сигнала окончания переноса (стоп-пептида). При чередовании в полипептиде сигналов начала переноса и окончания переноса белок будет пронизывать билипидный слой несколько раз. Синтезированные в шероховатом ЭПР белки подвергаются обработке. Наряду с отрезанием сигнального пептида, важнейшим преобразованием является гликозилирование (соединение с олигосахаридом). Здесь же происходят начальные этапы преобразования олигосахаридов в гликопротеинах. В цистернах гранулярного ЭПР обеспечивается также и правильное сворачивание синтезированных белковых молекул (гидрофобные участки ориентированы внутрь). Это препятствует образованию агрегатов, выпадающих в осадок. В гранулярном ЭПР происходит сборка липопротеиновых мембран. Здесь синтезируются не только мембранные белки, но и липиды мембран. Ферментативный синтез фосфолипидов происходит на обращенной к цитозолю стороне мембраны. Специальные белки-транслокаторы фосфолипидов могут переносить часть липидов во внутренний слой, создавая тем самым асимметрию билипидного слоя. ЭПР поставляет с помощью транспортных пузырьков мембранные белки и липиды аппарату Гольджи, который в свою очередь с помощью транспортных пузырьков снабжает ими плазмалемму и мембраны лизосом. 38. Строение аппарата Гольджи. Секреторная функция аппарата Гольджи. 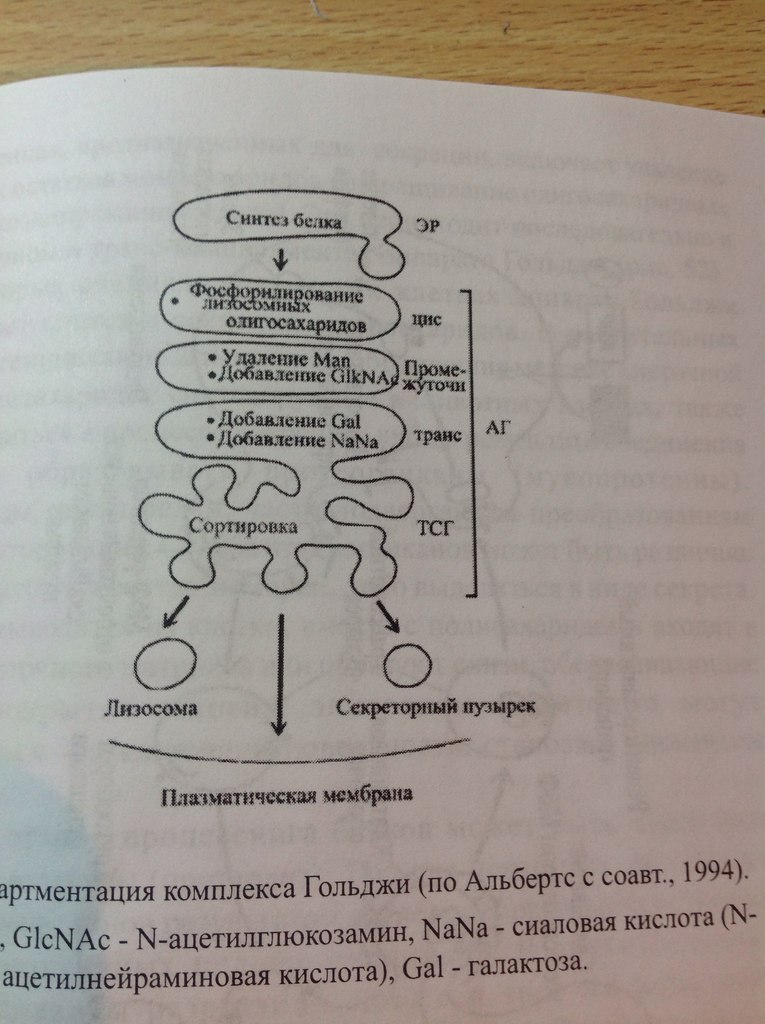 Аппарат Гольджи- интегрирующая часть метаболической системы эукариотической клетки. В нем происходит обособление секретируемых продуктов, идут процессы синтеза и модификации веществ с последующей их сортировкой. Он (АГ) осуществляет процессы секреции и снабжает гидролазами лизосомы. Основу АГ составляет диктиосома, которая включает стопку уплощенных цистерн, транспортные мембранные пузырьки, вакуоли и трубчатые структуры. АГ может быть представлен как единичной диктиосомой, так и целой системой диктиосом, которые расположены в разных участках цитоплазмы либо изолированно друг от друга, либо связаны между собой мембранными каналами. АГ обычно поляризован, что проявляется наличием цис- полюса (формирующаяся сторона) , промежуточной части и транс-полюса (зрелая сторона). Цистерны на цис-полюсе образуют цис-компартмент. Они, как правило, уплощенные и выпуклой поверхностью примыкают к ЭПР. Синтезированные в ЭПР вещества поступают в эти цистерны. На транс-полюсе цистерны образуют транс-компартмент. Эти цистерны расширены и заполнены секретом.к последней плоской цистерне транс-полюса примыкает транс-сеть аппарата Гольджи из трубчатых элементов и массы вакуолей. Вещества покидают АГ в составе мембранных пузырьков, образующихся на транс-полюсе и обеспечивающих их доставку. Секреция. Вещества выводятся из клетки одним из двух путей: в результате конструктивной (нерегулируемой) секреции (постоянная,например, путь мембранных белков через АГ) либо благодаря регулируемой(Факультативная, не постоянная, пример- путь внутриклеточных и секреторных белков) секреции. 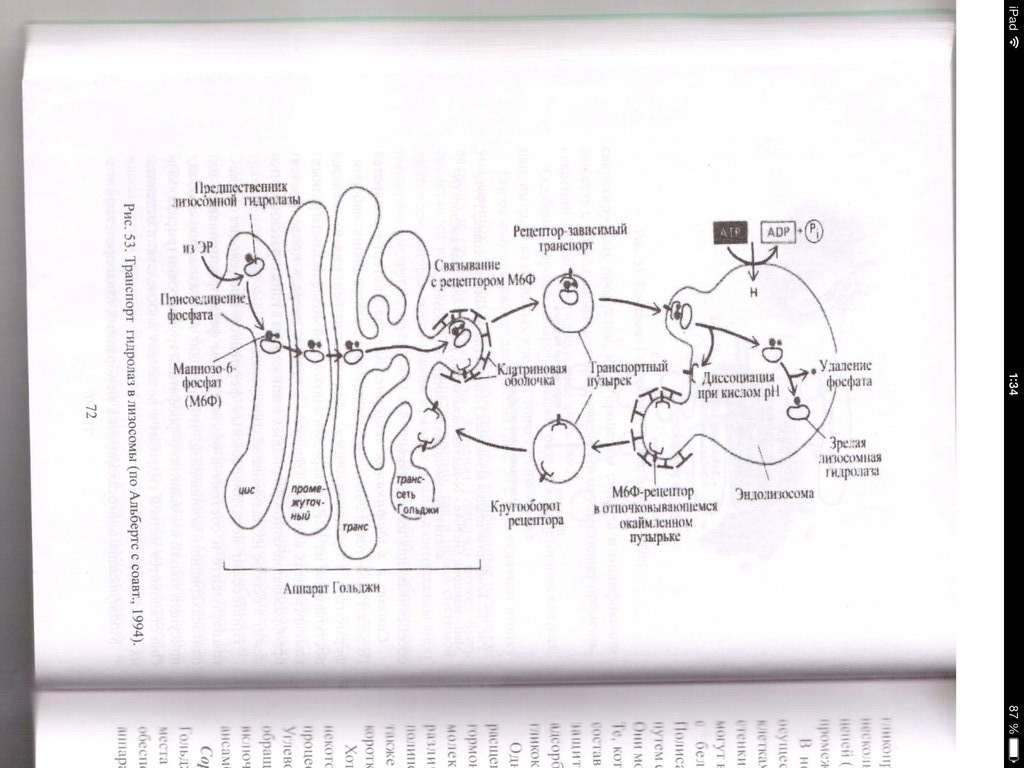 Механизм регулируемой секреции характерен для секреторных клеток. В этом случае белки избирательно упаковываются и отделяются от АГ в составе секреторных пузырьков. Секрет высвобождается из клетки только после сигнала (например, воздействия гормона или нейромедиатора). При конструктивной секреции, которая характерна для всех клеток, от АГ непрерывным потоком отделяются транспортные пузырьки, доставляющие секрет к плазмалемме. Таким образом доставляются к поверхности элементы гликокаликса, а также гликопротеины, протеогликаны, входящие в состав основного вещества соединительной ткани. 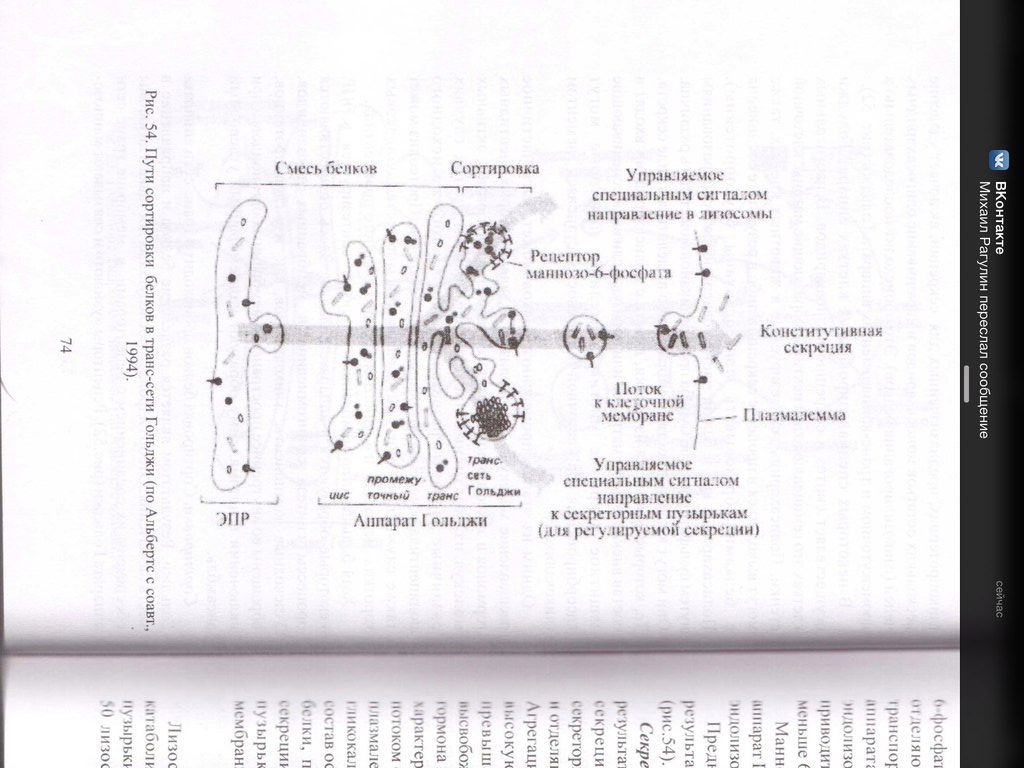 В некоторых специализированных клетках АГ осуществляется синтез и секреция полисахаридов. 39. Модификация белков в аппарате Гольджи. Сортировка белков в аппарате Гольджи. Сортировка белков происходит в транс-сети аппарата Гольджи. Результатом является разделение белков и направление в места назначения. Отбор гидролаз и последующая доставка в лизосомы обеспечены их соединением с рецептором в мембране транс-сети АГ. Рецепторы «узнают» и связывают маннозо-6-фосфатные метки лизосомных ферментов. В результате гидролазы отделяются от остальных белков и собираются в окаймленные транспортные пузырьки, которые отделяются от АГ, теряют свою кайму после чего сливаются с эндолизосомами или с лизосомами. Маннозо-6-фосфатные рецепторы возвращаются обратно в АГ в составе мембранных пузырьков, отрывающихся от эндолизосом. Предназначенные для секреции или для плазмалеммы белки в результате сортировки попадают в другие транспортные потоки. 40. Лизосомы. Образование, строение и функции. Морфологическая гетерогенность лизосом. Лизосомные патологии. 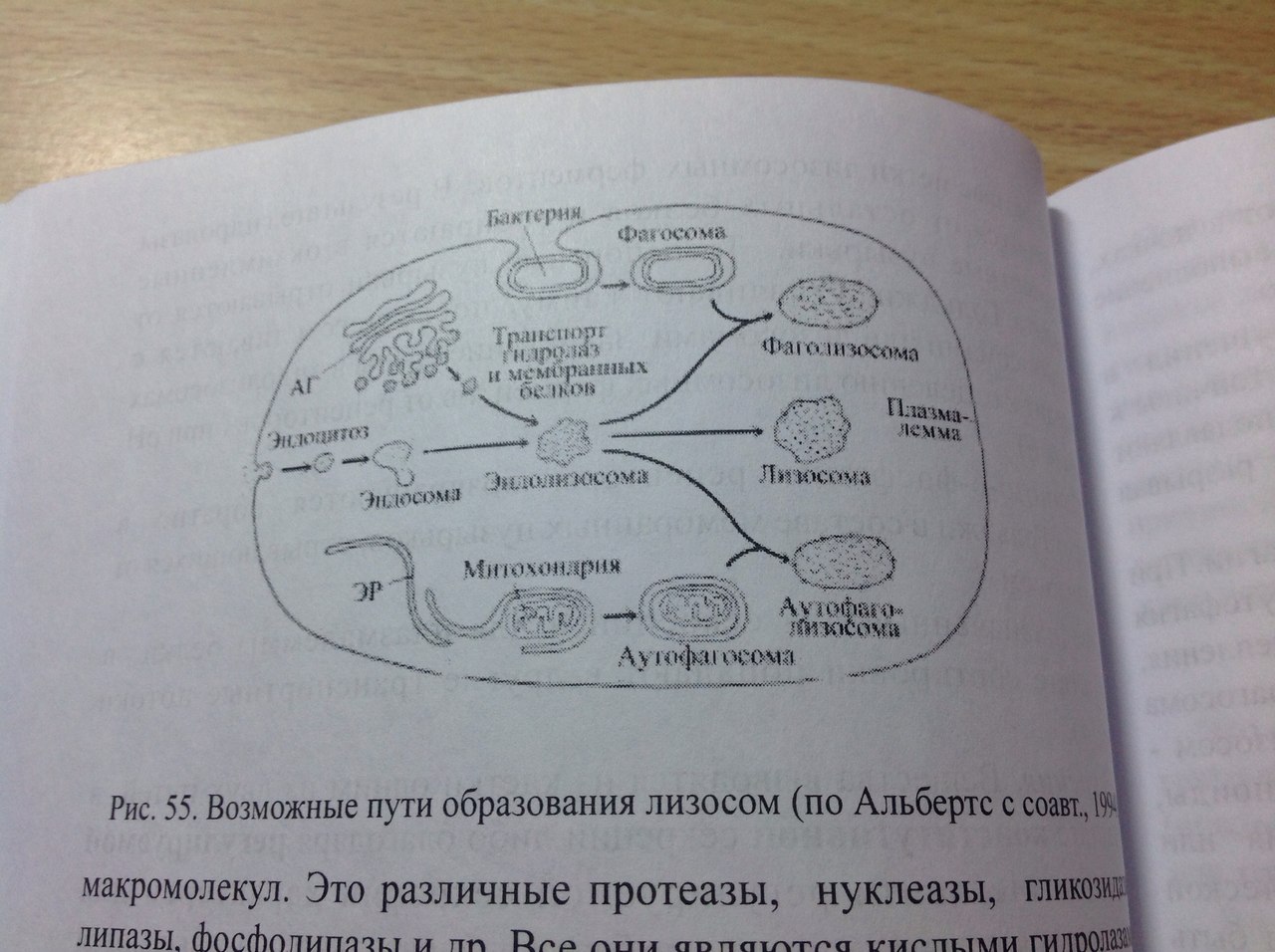 Лизосомы-специализированная часть внутриклеточной катаболической системы. Они представляют собой мемьранные пузырьки, содержащие ферменты. Известно более 50 лизосомных ферментов, которые обеспечивают расщепление макромолекул (различные протеазы, нуклеазы, гликозидазы, липазы…). Все они являются кислыми гидролазами. Мембрана лизосомы обладает рядом особенностей. Она образует барьер, устойчивый к действию гидролаз и предотвращающий их утечку. Мембрана содержит АТФ-зависимый протонный насос, который осуществляет закисление среды внутри лизосомы. Транспортные белки в мембране лизосомы переносят низкомолекулярные продукты расщепления в цитозоль. Первичная функция лизосом-участие в процессах внутриклеточного расщепления макромолекул до низкомолекулярных соединений. Поступление веществ в лизосомы происходит разными путями: 1.различные макромолекулы из внеклеточного пространства поступают в клетку в окаймленных пузырьках, образующихся в результате эндоцитоза. После этого они последовательно обнаруживаются сперва в периферических, затем в перинкулеарных эндосомах и наконец подвергаются действию гидролаз в эндолизосомах. Ферменты доставляются в эндолизосомы из аппарата Гольджти транспортными пузырьками, которые иногда называют первичными лизосомами. Дальнейшее расщепление происходит в зрелых лизосомах, называемых также вторичными. 2.при поглощении крупных объектов образуются фагосомы. Их слияние с эндолизосомами приводит к образованию фаголизосом. 3. характерен для аутофагии. Происходит расщепление собственных компонентов клетки. Аутофагия начинается с отделения органоида, предназначенного для расщепления от цитоплазмы за счет мембраны ЭПР. Получившаяся аутофагосома сливается с эндолизосомой и образуется аутофаголизосома. При аутофагии перевариваются органоиды, утратившие функциональную активность вследствие старения или регенерации отдельных клеточных структур. Она также может быть направлена на реализацию специальных задач (например, обеспечение жизнедеятельности при голодании). После накопления в лизосоме непереваренных остатков она превращается в остаточное тельце или телолизосому. Содержимое телолизосомы либо выводится путем экзоцитоза, либо длительное время хранится в клетке. В многоклеточных организмах гидролазы лизосом необходимы для гистогенеза и дифференцировки органов. Они обеспечивают процессы деструкции личиночных тканей при метаморфозе. В том случае имеет место автолиз-самопереваривание клеток в результате высвобождения ферментов лизосом. Он происходит также после гибели клетки и при некоторых патологиях. Многие наследственные заболевания связаны с нарушением работы лизосом (болезни накопления). Они могут быть вызваны дефектом гидролах\з. Это приводит к снижению активности ферментов и перегрузке лизосом непереваренными веществами. Например, при болезни Тея-Сакса нарушено расщепление липидов. Больные дети страдают задержкой умственного развития и умирают в возрасте 2-4 лет. Еще одной причиной развития болезней накопления может быть нарушение сортировки и транспорта ферментов в лизосомы, как в случае I-клеточной болезни. 41. Опишите путь секреторного белка от места синтеза до выхода из клетки. См. схема вопрос№39 Синтез секреторных белков завершается на гранулярном ЭПР. Секреторные белки, вместе с другими видами белков через транспортный пузырек попадает из гранулярного ЭПР в цис-полюс(незрелый) диктиосомы комплекса Гольджи . из цис-полюса транспортируется в промежуточную цистерну, оттуда дплее в транс-полюс, а затем в составе секреторных гранул переносятся к плазмалемме, откуда эндоцитозом попадают в межклеточное пространство. 42. Опишите путь гидролаз от места их синтеза до места назначения. Сборка любого белка начинается на свободных рибосомах в цитозоле. В ЭПР поступают только те пептиды, у которых первым синтезируется специфический гидрофобный сигнальный пептид. Особая частица, распознающая сигнал (SRP) связывается с сигнальным пептидом, временно блокирует синтез белка и затем направляет рибосому к мембране эндоплазматического ретикулума, где SRP присоединяется к своему рецептору. Доставленная таким образом к ЭПР рибосома прикрепляется своей большой субъединицей к специальному белку-рецептору, который участвует в образовании канала. Частица, выполнившая свою задачу, покидает рибосому. Прекращается ее блокирующее действие, и синтез белка возобновляется. Растущая белковая цепь поступает в ЭПР через канал в мембране. Пока белок в виде петли перетаскивается в полость ЭПР, его гидрофобный сигнальный пептид остается погруженным в мембрану. При синтезе растворимых белков сигнальный пептид отрезается, и белок высвобождается в полость ЭПР. На рибосомах гранулярного ЭПР происходит синтез белков, которые в зависимости от их конечного назначения могут быть разделены на три группы: 1.секреторные белки. 2.внутриклеточные белки(гидролазы) 3.мембранные белки. В ЭПР происходят начальные этапы сортировки синтезированных белков. Растворимые белки первых двух групп целиком поступают в цистерны ЭПР, который обеспечивает их обособления от цитозоля. Мембранные белки после синтеза остаются в составе мембраны ЭПР. 1)Отбор гидролаз и последующая доставка в лизосомы обеспечены их соединением с рецептором в мембране транс-сети АГ. 2)Рецепторы «узнают» и связывают маннозо-6-фосфатные метки лизосомных ферментов. 3)В результате гидролазы отделяются от остальных белков и собираются в окаймленные транспортные пузырьки, которые отделяются от АГ, теряют свою кайму после чего сливаются с эндолизосомами или с лизосомами. 4)Маннозо-6-фосфатные рецепторы возвращаются обратно в АГ в составе мембранных пузырьков, отрывающихся от эндолизосом. |