физиология экзамен. Физиология мои ответы экзамен. 40. Детектирование сигналов и опознание образов. Детектирование сигналов
 Скачать 4.25 Mb. Скачать 4.25 Mb.
|

|
48. Висцеросенсорная система, ее функции: - Интерорецепторы, их виды и механизмы возбуждения: Висцеросенсорная система воспринимает изменения внутренней среды организма и поставляет центральной и автономной нервной системе информацию, необходимую для рефлекторной регуляции работы всех внутренних органов. Интерорецеторы. Описаны разнообразные интерорецепторы, которые представлены свободными нервными, инкапсулированными нервными окончаниями: пластинчатые тельца (тельца Фатера—Пачини), колбы Краузе, расположенные на особых гломусных клетках. Механорецепторы реагируют на изменение давления в полых органах и сосудах, их растяжение и сжатие. Хеморецепторы сообщают ЦНС об изменениях химизма органов и тканей. Их роль особенно велика в рефлекторном регулировании и поддержании постоянства внутренней среды организма. Возбуждение хеморецепторов головного мозга может быть вызвано высвобождением из его элементов гистамина, индольных соединений, изменением содержания в желудочках мозга СО2 и другими факторами. Рецепторы каротидных клубочков реагируют на недостаток в крови кислорода, на снижение величины рН (в пределах 6,9— 7,6) и повышение напряжения СО2. Терморецепторы ответственны за начальный, афферентный этап процесса терморегуляции. Сравнительно мало исследованными остаются пока осморецепторы: они обнаружены в интерстициальной ткани вблизи капилляров. - проводниковый и корковый отделы висцеросенсорной системы: Проводящие пути сенсорной системы представлены, в основном, блуждающим, чревным и тазовым нервами. Блуждающий нерв передаёт афферентные сигналы в ЦНС по тонким волокнам с малой скоростью от практически всех органов грудной и брюшной полости, чревный нерв – от желудка, брыжейки и тонкого кишечника, а тазовый – от органов малого таза. В составе этих нервов имеются как быстро-, так и медленнопроводящие волокна. Импульсы от многих интероцепторов проходят по задним и вентролатеральным столбам спинного мозга. Корковым отделом висцеральной сенсорной системы являются чувствительные зоны коры больших полушарий, расположенные в основном в теменной доле в области Роландовой борозды. Однако в формировании условных рефлексов с участием интерорецепторов участвует лимбическая система (зона ствола мозга, отвечающая за вегетативные функции) и ассоциативные зоны коры большого мозга. 49. Болевая (ноцицептивная) сенсорная система, ее функции. Ноцицептивная (болевая) сенсорная система осуществляет формирование болевых ощущений, которые возникают при воздействии повреждающих факторов. В отличие от других сенсорных систем, не предоставляет информацию об окружающем мире. Однако она имеет не менее важное значение, поскольку информирует человека о предстоящей опасности, а, следовательно, играет защитную роль. Выделяют два вида боли: 1. Соматическую, возникающую при повреждении кожи и опорно-двигательного аппарата. Соматическую боль подразделяют на поверхностную и глубокую. Поверхностной болью называется боль кожного происхождения, а если ее источник локализован в мышцах, костях и суставах, она называется глубокой болью. Поверхностная боль проявляется в покалывании, пощипывании. Глубокая боль, как правило, тупая, плохо локализуется, обладает тенденцией иррадиировать в окружающие структуры, сопровождается неприятными ощущениями, тошнотой, сильным потоотделением, падением артериального давления. 2. Висцеральную, возникающую при повреждении внутренних органов и имеющую аналогичную картину с глубокой болью. - отраженная и проецированная боль, механизм их развития; Существуют особые виды боли – проекционная и отраженная. В качестве примера проекционной боли можно привести резкий удар по локтевому нерву. Подобный удар вызывает неприятное, трудно описываемое ощущение, распространяющееся на те участки руки, которые иннервируются этим нервом. Их возникновение основано на законе проекции боли: какая бы часть афферентного пути не раздражалась, боль ощущается в области рецепторов данного сенсорного пути. Одна из распространенных причин проекционных болей – это пережатие спинальных нервов в местах их вхождения в спинной мозг в результате повреждения межпозвонковых хрящевых дисков. Афферентные импульсы в ноцицептивных волокнах при такой патологии вызывают болевые ощущения, которые проецируются в область, связанную с травмируемым спинальным нервом. К проекционным (фантомным) болям относятся также боли, которые ощущают больные в области удаленной части конечности. Отраженными болями называются болевые ощущения не во внутренних органах, от которых поступают болевые сигналы, а в определенных частях кожной поверхности. Так, при стенокардии кроме болей в области сердца ощущается боль в левой руке и лопатке Отраженная боль отличается от проекционной боли тем, что она вызывается не прямой стимуляцией нервных волокон, а раздражением каких-либо рецептивных окончаний. Возникновение этих болей связано с тем, нейроны, проводящие болевые импульсы от рецепторов пораженного органа и рецепторов соответствующего участка кожи, конвергируют на одном и том же нейроне спиноталамического пути. Раздражение этого нейрона с рецепторов пораженного органа в соответствии с законом проекции боли приводит к тому, что боль ощущается и в области кожных рецепторов. - периферический, проводниковый и корковый отделы болевой сенсорной системы; Периферический отдел болевого анализатора. Представлен рецепторами боли, которые по предложению Ч. Шерлингтона называют ноцицепторами (от латинского слова «nocere» - разрушать). Эти высокопороговые рецепторы, реагирующие на раздражающее действие факторов. По механизму возбуждения ноцицепторы делят на механоноцицепторы и хемоноцицепторы. Механорецепторы расположены преимущественно в коже, фасциях, суставных сумках и слизистых оболочках пищеварительного тракта. Это свободные нервные окончания группы АΔ (дельта; скорость проведения 4 – 30 м/с). Реагируют на деформирующие воздействия, возникающие при растяжении или сжатии тканей. Большинство из них хорошо адаптируются. Хеморецепторы расположены также на коже и слизистых внутренних органов, в стенках мелких артерий. Представлены свободными нервными окончаниями группы С со скоростью проведения 0,4 – 2 м/с. Реагируют на химические вещества и воздействия, создающие дефицит О2 в тканях нарушающие процесс окисления (т.е. на алгогены). К таким веществам относятся: 1) тканевые алгогены – серотонин, гистамин, АХ и другие, образуются при разрушении тучных клеток соединительной ткани. 2) плазменные алгогены: брадикинин, простагландины. Выполняют функцию модуляторов, повышая чувствительность хемоноцицепторов. 3) Тахикинины при повреждающих воздействиях выделяются из окончаний нервов (вещество Р). Воздействуют местно на мембранные рецепторы того же нервного окончания. Проводниковый отдел. I нейрон – тело в чувствительном ганглии соответствующих нервов, иннервирующих определенные участки организма. II нейрон – в задних рогах спинного мозга. Далее болевая информация проводится двумя путями: специфическим (лемнисковым) и неспецифическим (экстралемнисковым). Специфический путь начинается от вставочных нейронов спинного мозга. В составе спиноталамического тракта импульсы поступают к специфическим ядрам таламуса, (III нейрон), аксоны III нейрона достигают коры. Неспецифический путь несет информацию от вставочного нейрона к различным структурам мозга. Выделяют три основных тракта, неоспиноталамический, спиноталамический и спиномезэнцефалический. Возбуждение по этим трактам поступает в неспецифические ядра таламуса, оттуда во все отделы коры больших полушарий. Корковый отдел. Специфический путь заканчивается в соматосенсорной зоне коры. Здесь происходит формирование острой, точно локализованной боли. Кроме того, за счет связей с моторной корой осуществляются моторные акты при воздействии болевых раздражений, происходит осознание и выработка программ поведения при болевом воздействии. Неспецифический путь проецируется в различные области коры. Особое значение имеет проекция в орбитофронтальную область коры, которая участвует в организации эмоционального и вегетативного компонентов боли. - теории происхождения боли: 1) Теория специфичности. Согласно этой теории формирование болевых ощущений возникает в следствие возбуждения специализированных рецепторов боли или ноцицепторов. Тем самым, появление импульсной активности в ноцицепторе сопровождается ощущением боли. 2) Теория интенсивности. Восприятие и ощущение боли формируется не только при возбуждении ноцицепторов, но и при любых возбуждениях рецепторов при условии, что раздражитель является сверхсильным. Тем самым, согласно теории интенсивности, болевые ощущения формируются зависимости от интенсивности раздражителя. 3) Паттерн-теория. Болевые ощущения формируются на основе специфических кодов и импульсной активности, поступающих по различным волокнами в цнс. Тем самым, формируется специфический паттерн импульсов, то есть определённый код, что приводит к формированию боли. 4) Теория «входных ворот». В области задних рогов спинного мозга имеются специализированные нейроны, образующие substantia gelatinosa. Эти нейроны выделяют медиаторы виде эндогенных опиоидов, а именно: эндорфины и энкефалины. В результате активация substantia gelatinosa приводит к блокаде поступления болевой информации, т.е. ворота закрываются, а подавление substantia gelatinosa приводит к открытию ворот и поступлению болевой информации. - Активация substantia gelatinosa чаще всего возникает при поступлении информации по волокнам типа А. В результате такой активации substantia gelatinosa блокирует поступление последующего импульсного потока к проекционному интернейрону в задних рогах спинного мозга, который является началом спиноталамического пути. На этом участке формируется пресимпатическое торможение и уменьшается выделение медиатора боли (например вещества Р). - Если же информация поступает по волокнам типа С, то substantia gelatinosa сама блокируется и тогда не происходит пресинаптического торможения и проекционный интернерон, возбуждаясь, передает информацию о боли по спинноталамическому тракту вышележащим структурам. Антиноцицептивная система, ее функции. Механизмы подавления боли: локальный контроль боли, нисходящий контроль боли. Антиноцицептивная система – противоболевая система. Контролирует порог болевой чувствительности и препятствует избыточности болевой информации. - Локальный контроль боли: Локальный контроль боли происходит на уровне задних рогов спинного мозга или чувствительных ядер черепно-мозговых нервов (например: 5 пары чмн). Суть локального контроля боли: на уровне задних рогов спинного мозга имеется substantia gelatinosa, которая препятствует поступлению болевой информации, выделяя медиаторы энкефалины, а также тормозные интернейроны, которые, выделяя ГАМК, препятствуют выделению ление болевого медиатора вещества Р, тем самым блокирует поток болевой информации. Таким образом, локальный контроль боли представлен теорией «входных ворот», поскольку речь идёт о substantia gelatinosa. Наряду с этим имеются тормозные клетки, вызывающие пресинаптическое торможение через ГАМК и, возможно, через рецептор. - Нисходящий контроль боли. Представлен нисходящими тормозными влияниями на проекционные интернейроны спинного мозга, которые являются началом спинаталамического пути, которым проводится болевая информация. На уровне ствола мозга к таким образованием относятся: ядра шва, голубого пятна, центральное серое околопроводное вещество среднего мозга. Эти образования выделяют ряд медиаторов, а именно: голубое пятно выделяет катехоламины, центральная серая околопроводное вещество – опиоидные вещества (эндорфины, энкефалины), а ядра шва – серотонин. Эти медиаторы отправляют свои влияния на задние рога спинного мозга и тем самым блокируют поток болевой импульсации. На уровне промежуточного мозга (таламус и гипоталамус) подлежащие нейросекреторные специализированные клетки также выделяют эндорфины и энкефалины. Кроме этого тормозные медиаторы в виде ГАМК, которые также своми нисходящими тормозными влияниями блокирует поток болевой импульсации. Таким образом, нисходящий контроль боли, за счёт нисходящих влияний вышележащих структур на спинной мозг и локальный контроль боли ограничивает поток болевой импульсации в ЦНС и препятствует формированию болевых ощущений. При длительном применении экзогенных наркотических веществ происходит подавление собственной антиноцицептивной системы, в частности, путём обратной связи уменьшается выделение эндорфинов и энкефалинов. В связи с чем возникает физическая зависимость. При этом, при отсутствии дополнительно введённых наркотических анальгетиков, у таких больных возникают выраженные болевые ощущения, даже при отсутствии сверхсильных раздражающих повреждающих факторов. 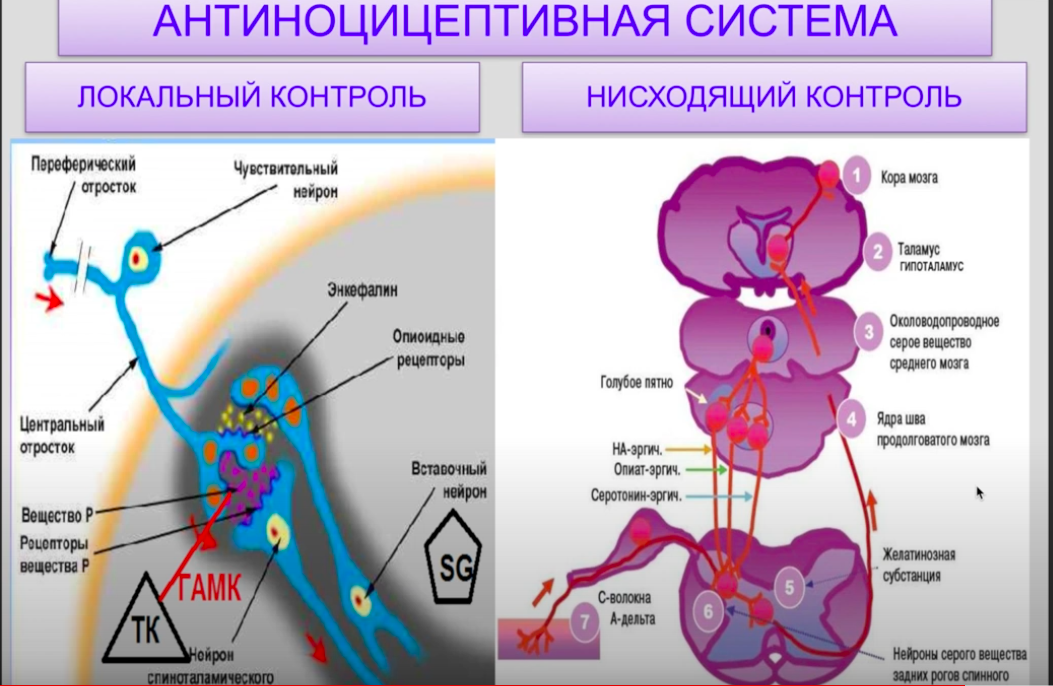 Вестибулярная сенсорная система, ее функции. - вестибулярный аппарат, строение и функции; механизм вестибулорецепии; Вестибулярная система обеспечивает восприятие и анализ информации о положении тела при угловых (вращательных) ускорениях, при прямолинейных движениях и поворотах головы, при воздействии сил гравитации. Периферический отдел вестибулярный системы расположен в перепончатом лабиринте и представлен рецепторным аппаратом, расположенным в трёх взаимноперпедикулярных полукружных каналах и преддверии улитки, а именно в эллиптическом мешочке ( или маточке) и в сферическом мешочке. Все эти структуры представлены эндолимфой, которая секретируется сосудистой полоской улитки и богата ионами калия. Полукружные каналы состоят из ножки и расширения ампулы. В ампуле содержится сенсорный эпителий гребешка с вестибулярными волосковыми клетками. Реснички этих клеток погружены в желеобразную структуру из мука полисахаридов - кубулу (с латинского – бочонок). При угловых вращательных ускорениях кубула отклоняется что приводит к сгибанию ресничек волосковых клеток. Каждый из полукружных каналов с одной стороны головы расположен в той же плоскости, что и соответствующий ему канал с другой стороны. Благодаря этому, участки сенсорного эпителия двух парных каналов воспринимают движение головы в любой плоскости. В эллиптическом мешочке (или маточке) и в сферическом мешочке сенсорной эпителий представлен в виде отолитового аппарата. В этом аппарате реснички волосковых клеток также, как реснички клеток ампулы, погружены в желеобразную мукополисахаридную массу. НО отличие этой массы состоит в том, что она содержит многочисленные отолиты или кристаллы карбоната кальция. Тем самым желеобразная масса с карбонатом кальция формирует отлитовую мембрану. Плотность данной мембраны в 2 раза выше, чем у эндолимфы, поэтому отолитовая мембрана легко двигается при прямолинейных ускорениях, в том числе создаваемых за счет сил гравитации. В эллиптическом мешочке (утрикулус) отолитовый аппарат ориентирован почти горизонтально, а в сферическом мешочке (саккулус) – вертикально. Поэтому прямолинейное ускорения могут быть восприняты как в горизонтальной плоскости (сгибание/разгибание головы), так и по вертикальной оси. Однако, угловые ускорения не приводит к движению отлитовой мембраны. Таким образом, рецепторы полукружных каналов возбуждаются при угловых, то есть вращательных движениях, а рецепторы отолитового аппарата при прямолинейных ускорениях, в том числе под действием сил гравитации. Волосковые клетки. Механизм возбуждения. Вестибулярные волосковые клетки делятся на два типа. Первый тип имеет форму колбы, второй тип - цилиндрической формы. Оба типа вестибулярных волосковых клеток образуют синаптические контакты с афферентными волокнами. Где афферентные волокна являются частью преддверно-улиткового нерва и тем самым обеспечивают передачу импульсации, возникающую на волосковых клетках. Также клетки первого и второго типа имеют эфферентную иннервацию. - Эфферентная иннервация для первого типа клеток характеризуется наличием синаптических контактов эфферентных волокон с афферентными волокнами. - Клетки второго типа характеризуются непосредственным синаптическим контактом волосковых клеток с афферентными волокнами. Естественно, что если афферентные волокна несут информацию к центрам вестибулярного анализатора, то эфферентные волокна характеризуются влиянием на чистоту импульсации волосковых клеток и тем самым обеспечивается нисходящей контроль вышележащих структур, разрядов клеток, а также происходит модуляции или коррекции частоты импульсивного потока по афферентным волокнам. Волосковые клетки характеризуются наличием следующих волосков: киноцилии и стереоцилии. Кинолиция одна и всегда располагается на периферии волосковых клеток. Стереоцилий больше и они располагаются по всей поверхности мембраны. Высота киноцилии превышает стереоцилии. Киноцилии и стереоцилии содержат актиновые филаменты. Они плотно взаимодействуют между собой и тем самым придают жёсткость киноцилию и стереоцилию. Считается, что подвижной является киноцилия, а стереоцилия более жёсткая. Однако у основания стереоцилий количество актиновых филаментов меньше, что позволяет своеобразному движению стереоцилию к киноцилию. Киноцилии и стереоцилии взаимодействуют между собой и связаны за счёт латеральных волокон, образованных различными белковыми компонентами. Наибольшую значимость имеют волокна эластического типа, которые обеспечивают связь между киноцилией и апикальным поверхностью стереоцилия. Именно это эластическое волокно является контролёром трансдукционных механоуправляемых каналов. Благодаря натяжению этих эластических волокон при движении киноцилий происходит открытие на апикальной поверхности мембраны трансдукционных каналов стереоцилий. Стереоцилии также взаимодействуют между собой за счет волокон и эти волокна, обладая эластическими свойствами обеспечивают открытие ионных каналов. |