физиология экзамен. Физиология мои ответы экзамен. 40. Детектирование сигналов и опознание образов. Детектирование сигналов
 Скачать 4.25 Mb. Скачать 4.25 Mb.
|

|
Механизм возбуждения волосковых клеток. Аналогичен общему принципу возбуждения, что характерно для вторичной чувствующих рецепторов. Но имеется ряд особенностей. Процесс возбуждения возникает вследствие движения эндолимфы. При этом движение эндолимфы направлено от стереоцилий к киноцилиям. В этом случае, ионные каналы открываются. Если же движение эндолимфы имеет противоположное направление, а именно от киноцилий к стереоцилиям натяжение эластических волокон уменьшается и ионные каналы на апикальной поверхности стереоцилий закрываются. Таким образом, при движении эндолимфы от стереоцилий к киноцилиям происходит натяжение эластических волокон, которые обеспечивают взаимодействие стереоцилий и киноцилий друг с другом и тем самым происходит открытие механотрансдукционных калиевых каналов, поскольку калия в эндолимфе больше. В результате открытия данных каналов калий поступает внутрь волосковой клетки, что приводит к развитию местной деполяризации (то есть рецепторного потенциала). Развитие рецепторного потенциала приводит к открытию кальцевых каналов засчет этого кальций поступает внутрь волосковой клетки, обеспечивает взаимодействие мембраны везикула с пресинаптической мембраной волосковой клетки, что сопровождается выделением медиатора путём экзоцитоза. Считается, что основным медиатором является ацетилхолин, но есть данные о наличии и глутамата. Тем самым, развитие рецепторного потенциала и выделение медиатора приводит к развитию уже потенциала действия (то есть генераторного потенциала) на первом нейроне вестибулярного анализатора. Если же движение эндолимфы имеет противоположное направление, то закрытие каналов сопровождается развитием гиперполяризации, что будет закрывать кальцевые каналы и выделение медиатора не произойдёт. 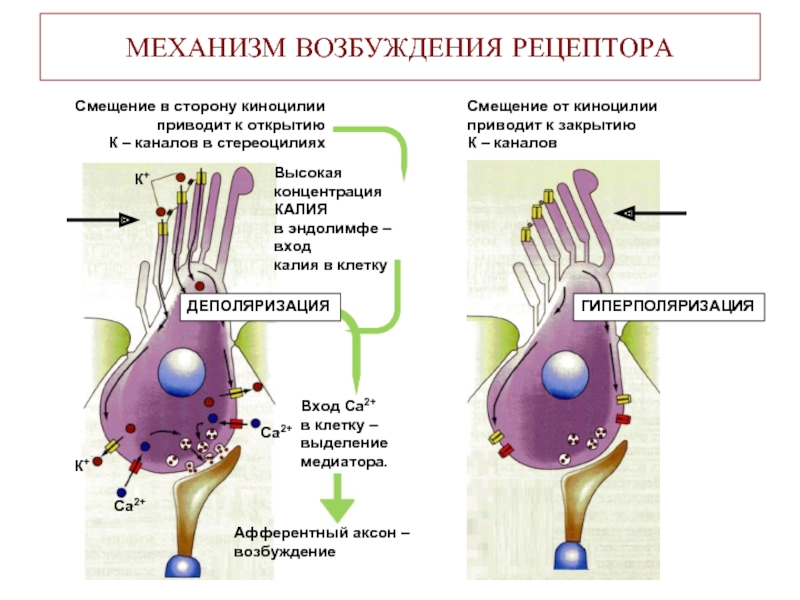 - проводниковый и корковый отделы вестибулярной сенсорной системы: Проводниковый отдел начинается 1 нейроном который расположен в вестибулярном ганглии. Аксоны этого нейрона формируют преддверную часть преддверно-улиткового нерва, т.е. 8 пары ЧМН. По аксонам 8 пары ЧМН импульсация поступает в ствол мозга к вестибулярным ядрам. Различают несколько вестибулярных ядер: верхние, нижние латеральные и медиальные ядра. Аксоны нейронов данных ядер несут информацию к специфическим ядрам таламуса, а от таламуса информация поступает в кору. Корковый отдел не имеет четкой локализации и представлен в теменной и в височной области неокортекса. - вестибулярные рефлексы, их характеристика; нистагм глаз: Вестибулярный анализатор имеет широкие связь со многими структурами ЦНС. В частности, у вестибулярных ядер имеются связи: - с мозжеском - с красными ядрами среднего мозга - с ядрами стриопаллидарной системы, что обеспечивает их роль в реализации поддержания равновесия и перераспределния тонуса мышц. - с центром регуляции висцеральных органов - с гипоталамусм - с сосудодвигательным центром продолговатого мозга - с центром рвоты За счет данных связей формируются вестибулярные рефлексы. К ним относят: 1) Вестибулоспинальный рефлекс. Формируется за счет связи вестибулярных ядер мотонейронами спинного мозга. За счет этого происходит перераспределение тонуса мышц туловища и обеспечивается поддержание равновесия при угловых и прямолинейных ускорениях. 2) При реализации поддержания равновесия играет связь вестибулярных ядер с мозжечком, базальными ядрами и другими компонентами ЦНС, которые в целом участвуют в регуляции движений. Связь вестибулярных ядер с центрами висцеральных функций приводит к формированию 2 группы рефлексов: Вестибуловегетативные рефлексы: - изменение частоты сокращений сердца - изменение тонуса сосудов при угловых и вращательных движениях - формирование чувства тошноты или возможной рвоты при изменении угловых ускорений. 3) Вестибулярные ядра имеют связь с двигательными ядрами тех черепно-мозговых нервов, которые иннервируют глазодвигательные мышцы, т.е. имеются связи с двигательными ядрами глазодвигательного, бокового и отводящего нервов. Благодаря данным связям формируются глазодвигательные рефлексы или нистагм глаз. Нистагм глаз – непроизвольное, ритмичное, скачкообразные двухкомпонентное движение глазных яблок. Различают быстрый и медленный компоненты. О направлении нистагма судят по быстрому компоненту. Однако, согласно закону Эвальда: направление движения эндолимфы соответствует именно медленный компонент. В зависимости от положения голову по отношению к туловищу во время вращения происходит движение эндолимфы в разных полукружных каналах, в связи с этим могут возникнуть разные виды нистагма: горизонтальный и вертикальный. Также существуют: оптокинетический нистагм (слежение за быстро перемещающимися объектами) установочный нистагм (при крайней позиции взгляда) Наличие спонтанного нистагма или усиление нистагма при отсутствии раздражения вестибулярного анализатора свидетельствует о наличии патологии. -------- ------- Морфофункциональная организация спинного мозга. Спинной мозг – наиболее древнее образование ЦНС позвоночных. Впервые появился у ланцетника. Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде передних корешков. Спинной мозг человека имеет 31-33 сегмента: 8 шейных (C1-C8), 12 грудных (Th1-Th11), 5 поясничных (L1-L5), 5 крестцовых (S1-S5), 1-3 копчиковых (Co1 -Co3). Морфологических границ между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным и определяется зоной распределения в нем волокон заднего корешка и зоной клеток, которые образуют выход передних корешков. Каждый сегмент через свои корешки иннервирует три метамера тела и получает информацию также от трех метамеров тела. В итоге перекрытия каждый метамер тела иннервируется тремя сегментами и передает сигналы в три сегмента спинного мозга. Спинной мозг человека имеет два утолщения: шейное и поясничное - в них содержится большее число нейронов, чем в других его участках. Волокна, поступающие по задним корешкам спинного мозга, выполняют функции, которые определяются тем, где и на каких нейронах заканчиваются данные волокна. В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние - эфферентными, двигательными, центробежными (закон Белла-Мажанди). Афферентные входы в спинной мозг организованы аксонами спинальных ганглиев, лежащих вне спинного мозга, и аксонами экстра- и интрамуральных ганглиев симпатического и парасимпатического отделов автономной нервной системы. Первая группа афферентных входов спинного мозга образована чувствительными волокнами, идущими от мышечных рецепторов, рецепторов сухожилий, надкостницы, оболочек суставов. Эта группа рецепторов образует начало так называемой проприоцептивной чувствительности. Проприоцептивные волокна по толщине и скорости проведения возбуждения делятся на 3 группы: 1) аннулоспиральные мышечные веретена – толщиной 12-20 мкм, скорость проведения возбуждения 110-120 м/с.; 2) вторичные окончания мышечных веретен – толщина волокон – 4-12 мкм, скорость проведения возбуждения – 35-70 м/с.; 3) пастинчатые тельца ( Фаттера – Пачини) - толщина волокон 1-4 мкм, скорость проведения возбуждения – 10 – 24 м/с. Волокна каждой группы имеют свои пороги возникновения возбуждения. Вторая группа афферентных входов спинного мозга начинается от кожных рецепторов: болевых, температурных, тактильных, давления - и представляет собой кожную рецептирующую систему. Третья группа афферентных входов спинного мозга представлена рецептирующими входами от висцеральных органов; это висцерорецептивная система. Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, и их волокна иннервируют все скелетные мышцы. 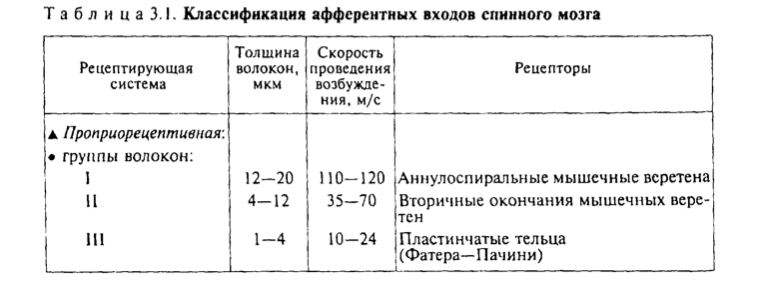  Восходящие пути (таблица ниже за два вопроса) Нисходящие проводящие пути спинного мозга, их функции. 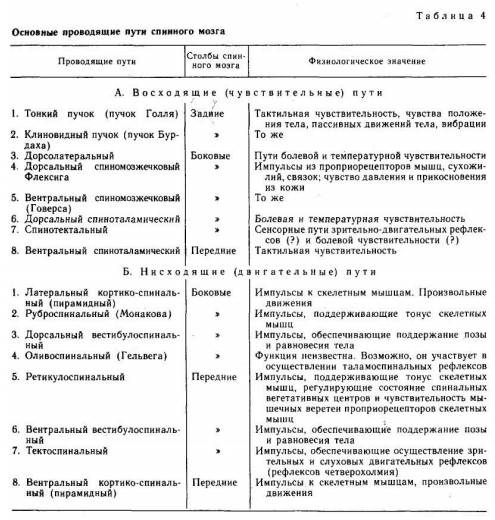 Нейроны сегментов спинного мозга. Функции задних и передних корешков сегментов спинного мозга (закон Белла и Мажанди). В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов. В связи с этим Шеррингтон назвал их «общим конечным путем». Эфферентные (двигательные) нейроны расположены в передних рогах спинного мозга, и их волокна иннервируют всю скелетную мускулатуру. Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом. Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга. Нейроны парасимпатического отдела автономной системы локализуются в сакральном отделе спинного мозга и являются фоновоактивными. В случаях раздражения и поражения задних корешков спинного мозга наблюдаются «стреляющие», опоясывающие боли на уровне метамера пораженного сегмента, снижение чувствительности всех видов, утрата или снижение рефлексов, вызываемых с метамера тела, который передает информацию в пораженный корешок. В случаях изолированного поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх. Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность. В опытах с перерезкой и раздражением корешков спинного мозга показано, что задние корешки являются афферентными, чувствительными, центростремительными, а передние — эфферентными, двигательными, центробежными (закон Белла—Мажанди). Функции нейронов боковых рогов сегментов спинного мозга. В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1—2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга — эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга. Средняя часть серого вещества спинного мозга содержит преимущественно короткоаксонные веретенообразные клетки (промежуточные нейроны), выполняющие связующую функцию между симметричными отделами сегмента, между клетками его передних и задних рогов. В случае поражения боковых рогов спинного мозга исчезают кожные сосудистые рефлексы, нарушается потоотделение, наблюдаются трофические изменения кожи, ногтей. При одностороннем поражении парасимпатического отдела автономной нервной системы на уровне крестцовых отделов спинного мозга нарушений дефекации и мочеиспускания не наблюдается, так как корковая иннервация этих центров является двусторонней. Альфа- и гамма-мотонейроны спинного мозга, их функции. Своими окончаниями аксон мотонейрона иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Чем меньше мышечных волокон иннервирует один аксон (т. е. чем меньше количественно мотонейронная единица), тем более дифференцированные, точные движения выполняет мышца. Мотонейронный пул – способность нескольких мотонейронов иннервировать одну мышцу. Возбудимость мотонейронов одного пула различна, поэтому при разной интенсивности раздражения в сокращение вовлекается разное количество волокон одной мышцы. При оптимальной силе раздражения сокращаются все волокна данной мышцы, таком случае развивается максимальное сокращение мышцы. Мотонейроны спинного мозга функционально делят на альфа- и гамма-нейроны. Альфа – мотонейроны имеют до 20 000 синапсов на своих дендритах и характеризуются низкой частотой импульсации (10—20 в секунду). Данные мотонейроны образуют прямые связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена. Гамма - мотонейроны имеют высокую частоту импульсации (до 200 в секунду) Данные нейроны получают информацию о его состоянии через промежуточные нейроны, иннервируя интрафузальные мышечные волокна мышечного веретена. Сокращение интрафузального мышечного волокна не приводит к сокращению мышцы, но повышает частоту разрядов импульсов, которые идут от рецепторов волокна в спинной мозг. Классификация спинномозговых рефлексов. Спинальный шок, его природа. С лекции Ашота ( видео цнс 1 часть примерно 7-8 минута) 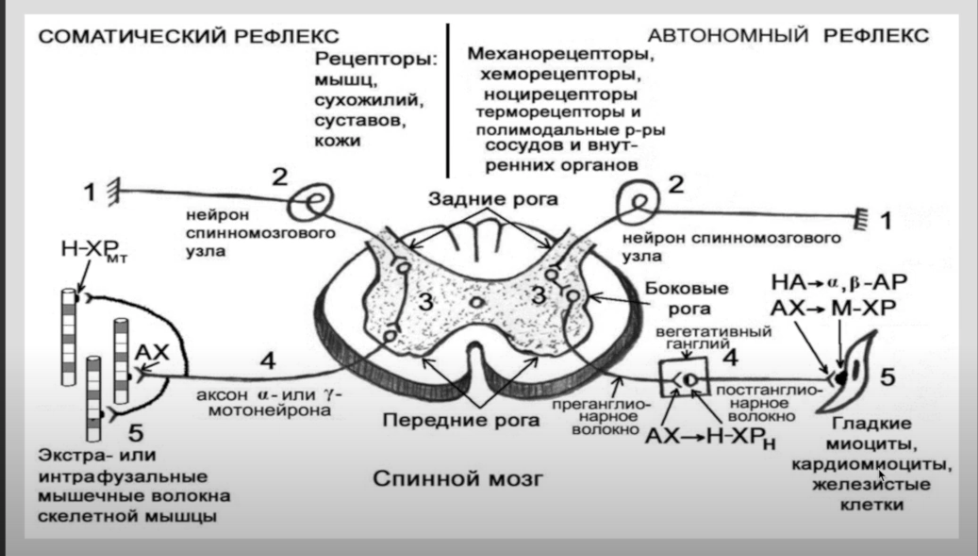 1) Соматосенсорные: рефлексы с рецепторов кожи носят характер, зависящий от силы раздражения, вида раздражаемого рецептора, но чаще всего конечная реакция выглядит в виде усиления сокращения мышц-сгибателей. 2) Соматовисцеральные: имеют свои пути; они начинаются от различных рецепторов, входят в спинной мозг через задние корешки, задние рога, далее в боковые рога, нейроны которых через передний корешок посылают аксоны не непосредственно к органам, а к ганглию симпатического или парасимпатического отдела автономной нервной системы; автономные (вегетативные) рефлексы обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов; эти рефлексы отличаются большим латентным периодом (ЛП) двумя фазами реакции: первая — ранняя — возникает с ЛП 7—9 мс и реализуется ограниченным числом сегментов, вторая — поздняя — возникает с большим ЛП — до 21 мс и вовлекает в реакцию практически все сегменты спинного мозга; поздний компонент вегетативного рефлекса обусловлен вовлечением в него вегетативных центров головного мозга; сложной формой рефлекторной деятельности спинного мозга является рефлекс, реализующий произвольное движение; в основе реализации произвольного движения лежит γ-афферентная рефлекторная система. В нее входят пирамидная кора, экстрапирамидная система, α- и γ-мотонейроны спинного мозга, экстра- и интрафузальные волокна мышечного веретена. 3) Висцеросоматические: рефлексы возникают при стимуляции афферентных нервов внутренних органов и характеризуются появлением двигательных реакций мышц грудной клетки и брюшной стенки, мышц разгибателей спины. 4) Миотатические рефлексы: рефлексы на растяжение мышцы; быстрое растяжение мышцы, всего на несколько миллиметров механическим ударом по ее сухожилию приводит к сокращению всей мышцы и двигательной реакции; например, легкий удар по сухожилию надколенной чашечки вызывает сокращение мышц бедра и разгибание голени; дуга этого рефлекса следующая: мышечные рецепторы четырехглавой мышцы бедра —> спинальный ганглий — > задние корешки —> задние рога III поясничного сегмента —> мотонейроны передних рогов того же сегмента —> экстрафузальные волокна четырехглавой мышцы бедра; реализация этого рефлекса была бы невозможна, если бы одновременно с сокращением мышц-разгибателей не расслаблялись мышцы- сгибатели; рефлекс на растяжение свойствен всем мышцам, но у мышц- разгибателей, они хорошо выражены и легко вызываются. Спинальный шок: Когда спинной мозг внезапно пересекается в верхней части шеи, сначала практически все функции спинного мозга, включая спинномозговые рефлексы, мгновенно подавляются, вплоть до полного их выключения. Эту реакцию называют спинальным шоком. Причиной такой реакции является то, что нормальная активность спинальных нейронов зависит в большой степени от постоянного тонического возбуждения спинного мозга под действием импульсов, поступающих к нему по нисходящим нервным волокнам от высших центров, особенно по ретикулоспинальным, вестибулоспинальным и кортикоспинальным трактам. В течение нескольких часов или недель возбудимость спинальных нейронов постепенно восстанавливается. По-видимому, это общее естественное свойство нейронов повсюду в нервной системе, т.е. после потери источника облегчающих импульсов нейроны увеличивают собственную естественную степень возбудимости, чтобы компенсировать потерю хотя бы частично. Большинству неприматов, чтобы возвратить к норме возбудимость центров спинного мозга, требуется от нескольких часов до нескольких суток. Однако у человека восстановление часто задерживается на несколько недель, а иногда полного восстановления совсем не происходит. В других случаях, наоборот, происходит чрезмерное восстановление с результирующей повышенной возбудимостью некоторых или всех функций спинного мозга. 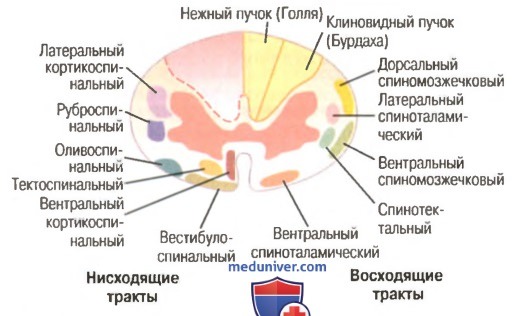 Поперечное сечение спинного мозга. Справа показаны главные восходящие тракты, слева — главные нисходящие тракты Далее перечислены некоторые из спинальных функций, особенно страдающих во время или после спинального шока. 1. В начале спинального шока сразу и очень значительно падает артериальное давление, иногда опускаясь ниже 40 мм рт. ст., что свидетельствует о практически полной блокаде активности симпатической нервной системы. Давление обычно возвращается к норме в течение нескольких дней (даже у человека). 2. Все рефлексы скелетных мышц, интегрированные в спинном мозге, во время первых стадий шока блокируются. Животным для восстановления этих рефлексов до нормы требуется от нескольких часов до нескольких дней; людям — от 2 нед до нескольких месяцев. И у животных, и у человека некоторые рефлексы могут в итоге стать чрезмерно возбудимыми, особенно в тех случаях, когда на фоне пересечения основной части путей между головным и спинным мозгом некоторые облегчающие пути сохраняются. Первыми восстанавливаются рефлексы на растяжение, в дальнейшем постепенно восстанавливаются более сложные рефлексы в соответствующем порядке: сгибательные, антигравитационные позные и частично шагательные. 3. Рефлексы крестцового отдела спинного мозга, контролирующие опорожнение мочевого пузыря и прямой кишки, подавлены у человека в течение первых недель после пересечения спинного мозга, но в большинстве случаев они в итоге восстанавливаются. |