физиология экзамен. Физиология мои ответы экзамен. 40. Детектирование сигналов и опознание образов. Детектирование сигналов
 Скачать 4.25 Mb. Скачать 4.25 Mb.
|

|
Неокортекс имеет шестислойное строение. 1) Первый слой — молекулярный. Содержит мало клеток, в основном — нервные волокна, образованные восходящими дендритами пирамидных нейронов и волокнами неспецифических ядер зрительного бугра, регулирующих уровень возбудимость коры больших полушарий. 2) Второй слой — наружный зернистый. Состоит из звездчатых клеток, ответственных за возбуждение в коре, т.е. за кратковременную память. 3) Третий слой — наружный пирамидный. Состоит из малых пирамидных клеток и совместно со 2-ым слоем обеспечивает «межкорковую» передачу информации. 4) Четвертый слой — внутренний зернистый. Содержит звездчатые клетки, на них заканчиваются специфические таламо-кортикальные афферентные пути от анализаторов. 5) Пятый слой — внутренний слой крупных пирамидных клеток. Их аксоны спускаются в продолговатый и спинной мозг. В моторной коре находятся гигантские пирамидные клетки Беца, чьи апикальные дендриты достигают поверхностных слоев, а самые длинные — образуют пирамидный тракт, достигающий спинного мозга. 6) Шестой слой — полиморфные клетки веретенообразной и треугольной формы. Образуют кортикоталамические пути. Сенсорные области: Корковые концы анализаторов имеют свою топографию и на них проецируются определенные афференты проводящих систем. Корковые концы анализаторов разных сенсорных систем перекрываются. Помимо этого, в каждой сенсорной системе коры имеются полисенсорные нейроны, которые реагируют не только на «свой» адекватный стимул, но и на сигналы других сенсорных систем. Кожная рецептирующая система, таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой извилины проецируются рецептивные поля кожи нижних конечностей, на средние — туловища, на нижние отделы — руки, головы. На заднюю центральную извилину в основном проецируются болевая и температурная чувствительность. В коре теменной доли (поля 5 и 7), где также оканчиваются проводящие пути чувствительности, осуществляется более сложный анализ: локализация раздражения, дискриминация, стереогноз. При повреждениях коры более грубо страдают функции дистальных отделов конечностей, особенно рук. Зрительная система представлена в затылочной доле мозга: поля 17, 18, 19. Центральный зрительный путь заканчивается в поле 17; он информирует о наличии и интенсивности зрительного сигнала. В полях 18 и 19 анализируются цвет, форма, размеры, качества предметов. Поражение поля 19 коры большого мозга приводит к тому, что больной видит, но не узнает предмет (зрительная агнозия, при этом утрачивается также цветовая память). Слуховая система проецируется в поперечных височных извилинах (извилины Гешля), в глубине задних отделов латеральной (сильвиевой) борозды (поля 41, 42, 52). Именно здесь заканчиваются аксоны задних бугров четверохолмий и латеральных коленчатых тел. Обонятельная система проецируется в области переднего конца гиппокампальной извилины (поле 34). Кора этой области имеет не шести-, а трехслойное строение. При раздражении этой области отмечаются обонятельные галлюцинации, повреждение ее ведет к аносмии (потеря обоняния). Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры (поле 43). Предисловие: простите шо так много тут просто такое-то мясо, нигде нет структурированной информации без воды, поэтому держите области с учебника ((( Ассоциативные области: Все сенсорные проекционные зоны и моторная область коры занимают менее 20% поверхности коры большого мозга. Остальная кора составляет ассоциативную область. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. Считают, что в ассоциативных областях происходит ассоциация разносенсорной информации. В результате формируются сложные элементы сознания. Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях. Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10—12%. Повреждение поля 18 приводит к зрительной агнозии. Больной видит, обходит предметы, но не может их назвать. Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры. В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации. Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных двигательных поведенческих актов. Первой и наиболее характерной чертой ассоциативных областей коры является мультисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта. Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации. Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти. Речевая функция связана как с сенсорной, так и с двигательной системами. Корковый двигательный центр речи расположен в заднем отделе третьей лобной извилины (поле 44) чаще левого полушария и был описан вначале Даксом (1835), а затем Брока (1861). Слуховой центр речи расположен в первой височной извилине левого полушария (поле 22). Этот центр был описан Вернике (1874). Моторный и слуховой центры речи связаны между собой мощным пучком аксонов. Речевые функции, связанные с письменной речью, — чтение, письмо — регулируются ангулярной извилиной зрительной области коры левого полушария мозга (поле 39). При поражении моторного центра речи развивается моторная афазия; в этом случае больной понимает речь, но сам говорить не может. При поражении слухового центра речи больной может говорить, излагать устно свои мысли, но не понимает чужой речи, слух сохранен, но больной не узнает слов. Такое состояние называется сенсорной слуховой афазией. Больной часто много говорит (логорея), но речь его неправильная (аграмматизм), наблюдается замена слогов, слов (парафазии). Поражение зрительного центра речи приводит к невозможности чтения, письма. Изолированное нарушение письма — аграфия, возникает также в случае расстройства функции задних отделов второй лобной извилины левого полушария. В височной области расположено поле 37, которое отвечает за запоминание слов. Больные с поражениями этого поля не помнят названия предметов. Они напоминают забывчивых людей, которым необходимо подсказывать нужные слова. Больной, забыв название предмета, помнит его назначения, свойства, поэтому долго описывает их качества, рассказывает, что делают этим предметом, но назвать его не может. Например, вместо слова «галстук» больной, глядя на галстук, говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости». Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, т. е. нейроны, реагирующие на различные раздражения. Например, при повреждении поля 17 зрительной области его функцию могут выполнять поля 18 и 19. Кроме того, разные двигательные эффекты раздражения одного и того же двигательного пункта коры наблюдаются в зависимости от текущей моторной деятельности. Если операцию удаления одной из зон коры провести в раннем детском возрасте, когда распределение функций еще не жестко закреплено, функция утраченной области практически полностью восстанавливается, т. е. в коре имеются проявления механизмов динамической локализации функций, позволяющих компенсировать функционально и анатомически нарушенные структуры. Важной особенностью коры большого мозга является ее способность длительно сохранять следы возбуждения. Следовые процессы в спинном мозге после его раздражения сохраняются в течение секунды; в подкорково-стволовых отделах (в форме сложных двигательно-координаторных актов, доминантных установок, эмоциональных состояний) длятся часами; в коре мозга следовые процессы могут сохраняться по принципу обратной связи в течение всей жизни. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления базы знаний. Сохранение следов возбуждения в коре проявляется в колебаниях уровня ее возбудимости; эти циклы длятся в двигательной области коры 3—5 мин, в зрительной — 5—8 мин. Основные процессы, происходящие в коре, реализуются двумя состояниями: возбуждением и торможением. Эти состояния всегда реципрокны. Они возникают, например, в пределах двигательного анализатора, что всегда наблюдается при движениях; они могут возникать и между разными анализаторами. Тормозное влияние одного анализатора на другие обеспечивает сосредоточенность внимания на одном процессе. Реципрокные отношения активности очень часто наблюдаются в активности соседних нейронов. Отношение между возбуждением и торможением в коре проявляется в форме так называемого латерального торможения. При латеральном торможении вокруг зоны возбуждения формируется зона заторможенных нейронов (одновременная индукция) и она по протяженности, как правило, в два раза больше зоны возбуждения. Латеральное торможение обеспечивает контрастность восприятия, что в свою очередь позволяет идентифицировать воспринимаемый объект. Помимо латерального пространственного торможения, в нейронах коры после возбуждения всегда возникает торможение активности и наоборот, после торможения — возбуждение — так называемая последовательная индукция. В тех случаях, когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения по коре. Иррадиация может происходить от нейрона к нейрону, по системам ассоциативных волокон I слоя, при этом она имеет очень малую скорость — 0,5—2,0 м/с. В другом случае иррадиация возбуждения возможна за счет аксонных связей пирамидных клеток III слоя коры между соседними структурами, в том числе между разными анализаторами. Иррадиация возбуждения обеспечивает взаимоотношение состояний систем коры при организации условнорефлекторного и других форм поведения. Наряду с иррадиацией возбуждения, которое происходит за счет импульсной передачи активности, существует иррадиация состояния торможения по коре. Механизм иррадиации торможения заключается в переводе нейронов в тормозное состояние под влиянием импульсов, приходящих из возбужденных участков коры, например, из симметричных областей полушарий. Моторная область коры большого мозга. Впервые Фритч и Гитциг (1870) показали, что раздражение передней центральной извилины мозга (поле 4) вызывает двигательную реакцию. В то же время признано, что двигательная область является анализаторной. В передней центральной извилине зоны, раздражение которых вызывает движение, представлены по соматотопическому типу, но вверх ногами: в верхних отделах извилины — нижние конечности, в нижних — верхние. Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры. В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области. Двигательная область коры, как никакая другая, имеет большое количество связей с другими анализаторами, чем, видимо, и обусловлено наличие в ней значительного числа полисенсорных нейронов. 75. Биоэлектрическая активность головного мозга. Ритмы ЭЭГ, их природа. Оценка функционального состояния коры большого мозга человека является трудной и до настоящего времени нерешенной задачей. Одним из признаков, косвенно свидетельствующим о функциональном состоянии структур головного мозга, является регистрация в них колебаний электрических потенциалов. Каждый нейрон имеет заряд мембраны, который при активации уменьшается, а при торможении - чаще увеличивается, т.е. развивается гиперполяризация. Глия мозга также имеет заряд клеток мембран. Динамика заряда мембраны нейронов, глии, процессы, происходящие в синапсах, дендритах, аксонном холмике, в аксоне - все это постоянно изменяющиеся, разнообразные по интенсивности, скорости процессы, интегральные характеристикой которых зависят от функционального состояния нервной структуры и суммарно определяют ее электрические показатели. Если эти показатели регистрируются через микроэлектроды, то они отражают активность локального (до 100 мкм в диаметре) участка мозга и называются фокальной активностью. В случае, если электрод располагается в подкорковой структуре, регистрируемая через него активность называется субкортикограммой; если электрод находится в коре мозга - кортикограммой. Наконец, если электрод помещен на поверхность кожи головы, то регистрируется суммарная активность как коры, так и подкорковых структур. Это проявление активности называется электроэнцефалограммой (ЭЭГ) (рис. 3.6). Все виды активности мозга в динамике подвержены усилению и ослаблению и сопровождаются определенными ритмами электрических колебаний. У человека в покое при отсутствии внешних раздражении преобладают медленные ритмы изменения состояния коры мозга, что на ЭЭГ находит отражение в форме так называемого альфа-ритма, частота колебаний которого составляет 8-13 в 1 с, а амплитуда - приблизительно 50 мкВ. Переход человека к активной деятельности приводит к смене аль- фа-ритма на более быстрый бета-ритм, имеющий частоту колебаний 14-30 в с, амплитуда которых составляет 25 мкВ. Переход от состояния покоя к состоянию сосредоточенного внимания или ко сну сопровождается развитием более медленного тета-ритма (4-8 колебаний в і с) или дельта-ритма (0,5-3,5 колебаний в с). Амплитуда медленных ритмов составляет 100-300 мкВ (см. рис. 3.6). Когда на фоне покоя или другого состояния предъявляется новое быстрое нарастающее раздражение, на ЭЭГ регистрируются так называемые вызванные потенциалы (ВП) - синхронная реакция множества нейронов данной зоны коры. Латентный период, амплитуда ВП зависят от интенсивности наносимого раздражения. Компоненты ВП, количество и характер его колебаний зависят от адекватности стимула относительно зоны регистрации ВП. ВП может состоять только из первичного или из первичного и вторичного ответов. Первичные ответы представляют собой двухфазные позитивно-негативные колебания. Они регистрируются в первичных зонах коры анализатора и только при адекватном для данного анализатора стимуле. Например, зрительная стимуляция для первичной зрительной коры (поле 17) является адекватной. Первичные ответы характеризуются коротким латентным периодом (ЛП), двухфазностью колебания: вначале положительная, затем отрицательная. Первичный ответ формируется за счет кратковременной синхронизации активности близлежащих нейронов. Вторичные ответы более вариабельны по ЛП, длительности, амплитуде, чем первичные. Как правило, вторичные ответы чаще возникают на сигналы, имеющие определенную смысловую нагрузку, на адекватные для данного анализатора стимулы; они хорошо формируются при обучении. Сверхмедленные биоэлектрические явления. С поверхности мозга регистрируются также постоянный потенциал и сверхмедленные колебания потенциалов. Постоянный потенциал определяется уровнем поляризации прилежащих к электроду образований мозга. Определенный вклад в поляризацию коры головного мозга вносит гематоэнцефалический барьер. Изменения метаболизма этих образований приводят к сверхмедленным колебаниям потенциала с периодами колебаний в секундах, декасекундах и минутах. Потенциалы, отводимые с кожи головы, отражают постоянный потенциал больших полушарий головного мозга. 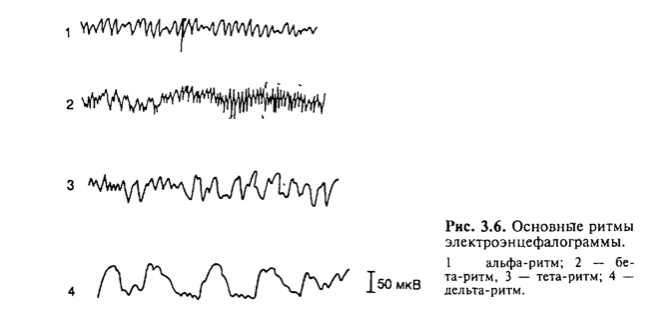 76. Межполушарные взаимоотношения. Межполушарная асимметрия. Взаимоотношение полушарий большого мозга определяется как функция, обеспечивающая специализацию полушарий, облегчение выполнения регуляторных процессов, повышение надежности управления деятельностью органов, систем органов и организма в целом. Роль взаимоотношений полушарий большого мозга наиболее четко проявляется при анализе функциональной межполушарной асимметрии. Асимметрия в функциях полушарий впервые была обнаружена в XIX в., когда обратили внимание на различные последствия повреждения левой и правой половины мозга. У некоторых больных при повреждении левого, а не правого полушария обнаруживались затруднения при чтении и письме. Считалось также, что левое полушарие управляет и «целенаправленными движениями». Совокупность этих данных стала основой представления о взаимоотношении двух полушарий. Одно полушарие (у праворуких обычно левое) рассматривалось как ведущее для речи и других высших функций, другое (правое), или «второстепенное», считали находящимся под контролем «доминантного» левого. Выявленная первой речевая асимметрия полушарий мозга предопределила представление об эквипотенциальности полушарий большого мозга детей до появления речи. Считается, что асимметрия мозга формируется при созревании мозолистого тела. Концепция доминантности полушарий, согласно которой во всех гностических и интеллектуальных функциях ведущим у «правшей» является левое полушарие, а правое оказывается «глухим и немым», просуществовала почти столетие. Однако постепенно накапливались свидетельства, что представление о правом полушарии как о второстепенном, зависимом, не соответствует действительности. Так, у больных с нарушениями левого полушария мозга хуже выполняются тесты на восприятие форм и оценку пространственных взаимосвязей, чем у здоровых. Неврологически здоровые испытуемые, владеющие двумя языками (английским и идиш), лучше идентифицируют английские слова, предъявленные в правом поле зрения, а слова на идиш — в левом. Был сделан вывод, что такого рода асимметрия связана с навыками чтения: английские слова читаются слева направо, а слова идиш — справа налево. Почти одновременно с распространением концепции доминантности полушарий стали появляться данные, указывающие на то, что правое, или второстепенное, полушарие также обладает своими особыми способностями. Так, Джексон выступил с утверждением о том, что в задних долях правого мозга локализована способность к формированию зрительных образов. Повреждение левого полушария приводит, как правило, к низким показателям по тестам на вербальные способности. В то же время больные с повреждением правого полушария обычно плохо выполняли невербальные тесты, включавшие манипуляции с геометрическими фигурами, сборку головоломок, восполнение недостающих частей рисунков или фигур и другие задачи, связанные с оценкой формы, расстояния и пространственных отношений. Обнаружено, что повреждение правого полушария часто сопровождалось глубокими нарушениями ориентации и сознания. Такие больные плохо ориентируются в пространстве, не в состоянии найти дорогу к дому, в котором прожили много лет. С повреждением правого полушария были связаны также определенные виды агнозий, т. е. нарушений в узнавании или восприятии знакомой информации, восприятии глубины и пространственных взаимоотношений. Одной из самых интересных форм агнозии является агнозия на лица. Больной с такой агнозией не способен узнать знакомого лица, а иногда вообще не может отличать людей друг от друга. Узнавание других ситуаций и объектов, например, может быть при этом не нарушено. Дополнительные сведения, указывающие на специализацию правого полушария, были получены при наблюдении за больными, страдающими тяжелыми нарушениями речи, у которых, однако, часто сохраняется способность к пению. Кроме того, в клинических сообщениях содержались данные о том, что повреждение правой половины мозга может привести к утрате музыкальных способностей, не затронув речевых. Это расстройство, называемое амузией, чаще всего отмечалось у профессиональных музыкантов, перенесших инсульт или другие повреждения мозга. После того как нейрохирурги осуществили серию операций с комиссуротомией и были выполнены психологические исследования на этих больных, стало ясно, что правое полушарие обладает собственными высшими гностическими функциями. Существует представление, что межполушарная асимметрия в решающей мере зависит от функционального уровня переработки информации. В этом случае решающее значение придается не характеру стимула, а особенностям гностической задачи, стоящей перед наблюдателем. Принято считать, что правое полушарие специализировано в переработке информации на образном функциональном уровне, левое — на категориальном. Применение такого подхода позволяет снять ряд трудноразрешимых противоречий. Так, преимущество левого полушария, обнаруженное при чтении нотных и пальцевых знаков, объясняется тем, что эти процессы протекают на категориальном уровне переработки информации. Сравнение слов без их лингвистического анализа успешнее осуществляется при их адресации правой гемисфере, поскольку для решения этих задач достаточна переработка информации на образном функциональном уровне. Межполушарная асимметрия зависит от функционального уровня переработки информации: левое полушарие обладает способностью к переработке информации как на семантическом, так и на перцептивном функциональных уровнях, возможности правого полушария ограничиваются перцептивным уровнем. В случаях латерального предъявления информации можно выделить три способа межполушарных взаимодействий, проявляющихся в процессах зрительного опознания. - Параллельная деятельность. Каждое полушарие перерабатывает информацию с использованием присущих ему механизмов. - Избирательная деятельность. Информация перерабатывается в «компетентном» полушарии. - Совместная деятельность. Оба полушария участвуют в переработке информации, последовательно играя ведущую роль на тех или иных этапах этого процесса. Основным фактором, определяющим участие того или иного полушария в процессах узнавания неполных изображений, является то, каких элементов лишено изображение, а именно какова степень значимости отсутствующих в изображении элементов. В случае, если детали изображения удалялись без учета степени их значимости, опознание в большей мере было затруднено у больных с поражениями структур правого полушария. Это дает основание считать правое полушарие ведущим в опознании таких изображений. Если же из изображения удалялся относительно небольшой, но высокозначимый участок, то опознание нарушалось в первую очередь при поражении структур левого полушария, что свидетельствует о преимущественном участии левой гемисферы в опознании подобных изображений. В правом полушарии осуществляется более полная оценка зрительных стимулов, тогда как в левом оцениваются наиболее существенные, значимые их признаки. Когда значительное число деталей изображения, подлежащего опознанию, удалено, вероятность того, что наиболее информативные, значимые его участки не подвергнутся искажению или удалению, невелика, а потому левополушарная стратегия опознания значительно ограничена. В таких случаях более адекватной является стратегия, свойственная правому полушарию, основанная на использовании всей содержащейся в изображении информации. Трудности в реализации левополушарной стратегии в этих условиях усугубляются еще и тем обстоятельством, что левое полушарие обладает недостаточными «способностями» к точной оценке отдельных элементов изображения. Об этом свидетельствуют также исследования, согласно которым оценка длины и ориентации линий, кривизны дуг, величины углов нарушается прежде всего при поражениях правого полушария. Иная картина отмечается в случаях, когда большая часть изображения удалена, но сохранен наиболее значимый, информативный его участок. В подобных ситуациях более адекватным является способ опознания, основанный на анализе наиболее значимых фрагментов изображения — стратегия, используемая левым полушарием. В процессе узнавания неполных изображений участвуют структуры как правого, так и левого полушария, причем степень участия каждого из них зависит от особенностей, предъявляемых изображений, и в первую очередь от того, содержит ли изображение наиболее значимые информативные элементы. При наличии этих элементов преобладающая роль принадлежит левому полушарию; при их удалении преимущественную роль в процессе опознания играет правое полушарие. |