физиология экзамен. Физиология мои ответы экзамен. 40. Детектирование сигналов и опознание образов. Детектирование сигналов
 Скачать 4.25 Mb. Скачать 4.25 Mb.
|

|
131. Эритропоэз, его нервная и гуморальная регуляция. Эритропоэз — процесс образования эритроцитов в костном мозге. Первой клеткой эритроидного ряда, образующейся из колониеобразующей клетки эритроцитарной (КОК-Э) — клетки-предшественницы эритроидного ряда, является проэритробласт, из которого в ходе 4—5 последующих удвоений и созревания образуется 16—32 зрелых эритроцита. Схема процесса: 1 проэритробласт (удвоение) => два базофильных эритробласта I порядка => 4 базофильных эритробласта II порядка => 8 полихроматофильных эритробластов I порядка => 16 полихроматофильных эритробластов II порядка => 32 полихроматофильных нормобласта => 32 оксифильных нормобласта => денуклеация нормобластов => 32 ретикулоцита => 32 эритроцита. Эритропоэз до формирования ретикулоцита занимает 5 дней. Эритропоэз у человека и животных (от проэритробласта до ретикулоцита) протекает в эритробластических островках костного мозга, которых в норме содержится до 137 на 1 мг ткани костного мозга. Макрофаги эритроцитарных островков играют основную роль в физиологии эритроидных клеток, влияя на их пролиферацию и созревание. Макрофаги фагоцитируют вытолкнутые из нормобластов ядра, обеспечивают эритробласты ферритином и пластическими веществами, секретируют эритропоэтин и гликозаминогликаны, последние повышают концентрацию ростковых факторов в островках. Эти благоприятные условия для развития эритробластов макрофаги создают благодаря наличию рецепторов к эритроидным клеткам-предшественницам. Из костного мозга ретикулоциты выходят в кровь и в течение суток созревают в эритроциты. По количеству ретикулоцитов в крови судят об эритроцитарной продукции костного мозга и интенсивности эритропоэза. У человека их количество составляет 5— 10%о. За сутки в 1 мкл крови поступает 60—80 тыс. эритроцитов. В 1 мкл крови у мужчин содержится 5,21 (4,52—5,9) млн, а у женщин — 4,6 (4,1—5,1) млн эритроцитов. Уменьшение количества эритроцитов в единице объема крови называется анемией, увеличение — эритроцитозом. Последний может носить физиологический, приспособленный для организма человека характер (например, при подъеме человека в горы, на высоту более 3000 м над уровнем моря). 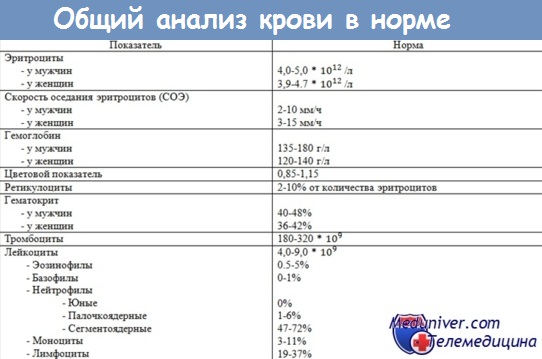 Из учебника: В норме клеточные элементы эритропоэза в костном мозге размножаются чрезвычайно интенсивно: за сутки в костном мозге образуется 2-1011 эритроидных клеток. При этом коммитированные эритроидные предшественники от момента образования из пСКК претерпевают от 5 до 10 делений, превращаясь вначале в бурстобразующую единицу эритроцитов - БОЕэ (burst - взрыв), а затем в колониеобразующую единицу эритроцитов (КОЕэ). Из КОЕэ, являющейся клеткой-предшественницей эритроидного ряда, вначале появляется эритробласт, который через ряд промежуточных стадии дает поколение юных эритроцитов - ретикулоцитов. Из одного эритробласта в результате митозов появляется от 16 до 32 ретикулоцитов. Ретикулоцит еще в течение суток находится в костном мозге, а затем поступает в кровоток, где за 50 -70 ч превращается в молодой эритроцит, или нормоцит. В крови здорового человека находится не более 1 % ретикулоцитов. Регуляция эритропоэза: Для нормального эритропоэза, кроме действия микроокружения, необходимо наличие микроэлементов, гормонов, витаминов, ростовых факторов, интерлейкинов и специфических факторов эритропоэза - эритропоэтинов. Железо. Для нормального эритропоэза в сутки требуется до 25 мг железа. Железо поступает в костный мозг в основном при разрушении эритроцитов. Взрослому человеку для нормального эритропоэза требуется в суточном рационе 12-15 мг железа. Всасыванию железа в кишечнике способствует аскорбиновая кислота, переводящая Fe3+ в Fe2+, который сохраняет растворимость при нейтральных и щелочных рН. На участке слизистой тонкой кишки имеются рецепторы, облегчающие переход железа в клетки тонкой кишки, а оттуда в плазму. В слизистой тонкой кишки находится переносчик железа – белок трансферрин. Он доставляет железо в клетки, имеющие трансферриновые рецепторы. В клетке комплекс трансферрина и железа распадается и Fe3+ соединяется с другим белком-переносчиком - ферритином. Железо в соединении с ферритином переносится непосредственно от клеток хранения к клеткам-предшественникам эритроцитов. Важным компонентом эритропоэза является медь, которая усваивается непосредственно в костном мозге, катализируя образование гемоглобина. Для эритропоэза необходимы и другие микроэлементы - кобальт, магний, марганец, селен, цинк и др. Для нормального эритропоэза требуются витамины, и в первую очередь витамин В12, и фолиевая кислота. Эти витамины оказывают сходное взаимодополняющее действие на эритропоэз. Для всасывания витамина В требуется внутренний фактор кроветворения - гастромукопротеин. Это комплексное соединение образуется в желудке. Немаловажную роль в регуляции эритропоэза играют другие витамины группы В, а также железы внутренней секреции. Все гормоны, регулирующие обмен белков (соматотропный и тиреотропный гормоны гипофиза, гормон щитовидной железы - тироксин и др.) и кальция (паратгормон, тиреокальцитонин), необходимы для нормального эритропоэза. Мужские половые гормоны (андрогены) слегка стимулируют эритропоэз, тогда как женские (эстрогены) тормозят его, что обусловливает меньшее число эритроцитов у женщин по сравнению с мужчинами. Цитокины. Особо важную роль в регуляции эритропоэза играют цитокины, и в первую очередь специфический регулятор эритропоэза эритропоэтин. Эритропоэтин является гликопротеидом с мол. массой 36 000. В небольших концентрациях эритропоэтины обнаружены в крови здоровых людей, что позволяет считать их физиологическими регуляторами эритропоэза. При анемиях, сопровождающих заболевания почек, эритропоэтины отсутствуют или их концентрация значительно снижается. Эти вещества синтезируются и секретируются в основном перитубулярными клетками почки. Эритропоэтин оказывает действие непосредственно на клетки-предшственники эритроидного ряда (БОЕэ и КОЕэ). Его функции сводятся к следующему: - ускоряет и усиливает переход БОЭэ в КОЕэ, а последние в эритробласты; - увеличивает число митозов клеток эритроидного ряда; - исключает один или несколько циклов митотических делений; - ускоряет созревание неделящихся клеток - нормобластов, ретикулоцитов; - увеличивает выход ретикулоцитов из костного мозга в общий кровоток; - усиливает синтез гемоглобина. После образования БОЕэ на нее, кроме эритропоэтина, оказывают влияние ИЛ-3 и GM-KSF, благодаря чему она превращается в КОЕэ, переходящую под воздействием эритропоэтина в эритробласт. 132. Количество и функции лейкоцитов. Лейкоцитарная формула. Возрастные изменения в лейкоцитарной формуле у детей. Лейкоциты (белые кровяные тельца) представляют собой образования различной формы и величины. По строению лейкоциты делятся на две группы: зернистые (гранулоциты) и незернистые (агранулоциты). К гранулоцитам относятся нейтрофилы, эозинофилы и базофилы, к агранулоцитам - лимфоциты и моноциты. В норме количество лейкоцитов у взрослых людей колеблется от 4,5 до 9 тыс. в 1 мм3, или 4,5-9*109/л. Увеличение числа лейкоцитов за пределы нормы называется лейкоцитозом, уменьшение - лейкопенией. Нейтрофилы в циркуляции живут от 8 до 10 ч. Находящиеся в кровотоке нейтрофилы могут быть условно разделены на 2 группы: 1) свободно циркулирующие и 2) нейтрофилы, занимающие краевое положение в сосудах. Между той и другой группой существует динамическое равновесие и постоянный обмен. Следовательно, в сосудистом русле нейтрофилов содержится приблизительно в 2 раза больше, чем определяется в вытекающей крови. Нейтрофилы - самая многочисленная популяция лейкоцитов. У взрослого человека ежедневно обменивается приблизительно 1,6-109 нейтрофилов, благодаря чему количество микрофагов сохраняется на постоянном уровне. Нейтрофилы постоянно мигрируют из сосудистого русла. Они подвергаются элиминации с секретами слизистых оболочек (особенно в полости рта) или в течение 2-6 дней погибают в тканях. Нейтрофилы содержат богатейший набор биологически активных субстанций, в том числе способных убивать бактерии, вирусы и раковые клетки. Нейтрофилы подвижны, легко проникают в экстравазальное пространство ткани, высокоактивны. При стимуляции нейтрофилы быстро реализуют свой цитолитический материал по тношению к вирусинфицированным и опухолевым клеткам и могут запускать у них генетические программы апоптоза. Нейтрофилы осуществляют цитотоксический эффект (киллинг) в отношении отдельных чужеродных клеток. Нейтрофил подходит к клетке-мишени и убивает ее на расстоянии с помощью активных форм кислорода, повреждающих мембрану. Нейтрофилы синтезируют и секретируют про-воспалительные цитокины - ФНОальфа, ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12, интерферон (Ифа), гранулоцитарномакрофагальный колониестимулирующий фактор (ГМ-КСФ), фактор, активирующий тромбоциты (ФАТ), фактор роста фибробластов В ответ на раздражители нейтрофилы активируются, проникают в поврежденные ткани, дегранулируют, высвобождают протеолитические и липолитические ферменты, обладающие бактерицидной активностью. Обладая фагоцитарной функцией, нейтрофилы поглощают не только бактерии, но и продукты повреждения тканей. Базофилы. В крови базофилов очень мало (40-60 в 1 мкл), однако в различных тканях, в том числе сосудистой стенке, содержатся тучные клетки, иначе называемые тканевыми базофилами, которые выполняют те же функции, что и базофилы. Базофилы в кровотоке живут часы, тогда как срок жизни тучных клеток исчисляется месяцами и даже годами. Функция базофилов и тучных клеток обусловлена наличием в них ряда биологически активных веществ. К ним в первую очередь принадлежит гистамин, расширяющий кровеносные сосуды. В базофилах содержатся противосвертывающие вещества - гепарин, хондроитинсульфаты А и С, дерматансульфат и гепарансульфат. Все перечисленные глюкозамингликаны в норме способствуют сохранению крови в жидком состоянии. Базофилы способны синтезировать и секретировать фактор, активирующий тромбоциты (ФАТ), - соединение, обладающее широким спектром действия, в частности резко усиливающее агрегацию тромбоцитов. Кроме того, базофилы синтезируют тромбоксаны (соединения, способствующие агрегации тромбоцитов), лейкотриены и простагландины. В базофилах и тучных клетках содержится ряд протеолитических ферментов, дегидрогеназы и пероксидаза, а также соединение, получившее наименование фактор хемотаксиса эозинофилов. Последний способствует привлечению эозинофилов из сосудов в места скопления базофилов в органах-мишенях. При этом эозинофилы поглощают гранулы базофилов и приводят к разрушению гистамина с помощью фермента гистаминазы. При сенсибилизации базофилы начинают продуцировать и секретировать нейтрофильный хемотаксический фактор и серотонин. Эозинофилы. Длительность пребывания эозинофилов в кровотоке не превышает 12 ч, после чего они проникают в ткани, где живут 10-12 сут. Эозинофилы содержат большое число гранул, в которых находятся ферменты и многие биологически активные соединения. Основным компонентом гранул является главный щелочной белок, имеющий мол. массу около 1000 и играющий важную роль в защите от паразитов. Его особенность заключается в том, что он способен нейтрализовать ряд ферментов - Бэта-глюкуронидазу, рибонуклеазу, фосфолипазу, а также медиаторы воспаления и гепарин. Все эти реакции необходимы для ликвидации последствий аллергических реакций в организме. В гранулах эозинофилов находятся гистаминаза, коллагеназа, эластаза, глюкуронидаза, катепсин, РНКаза, миелопероксидаза, кислая фосфатаза и арилсульфатаза В. Два последних фермента способны инактивировать анафилаксин, выделяемый при дегрануляции базофилами. Кроме того, эозинофилы секретируют простагландины. Эозинофилы обладают фагоцитарной активностью. Особенно интенсивно они фагоцитируют кокки. В тканях эозинофилы скапливаются преимущественно в тех органах, где содержится гистамин - в слизистой оболочке и подслизистой основе желудка и тонкй кишки, в легких. Здесь их число превышает содержание в крови в 200-300 раз. Эозинофилы захватывают гистамин и разрушают его с помощью фермента гистаминазы. В составе эозинофилов находится фактор, тормозящий выделение гистамина тучными клетками и базофилами. Эозинофилы играют важную роль в разрушении токсинов белкового происхождения, чужеродных белков и иммунных комплексов. Велика роль эозинофилов, осуществляющих цитотоксический эффект, в борьбе с гельминтами, их яйцами и личинками. В частности, при контакте активированного эозинофила с личинками происходит его дегрануляция с последующим выделением большого количества белка и ферментов на поверхность личинки, что приводит к разрушению последней. Увеличение числа эозинофилов, наблюдаемое при миграции личинок, является одним из важнейших механизмов в ликвидации гельминтозов. Содержание эозинофилов резко возрастает при аллергических заболеваниях, когда происходит дегрануляция базофилов и выделение анафилактического хемотаксического фактора, который привлекает эозинофилы. При этом эозинофилы выполняют роль «чистильщиков», фагоцитируя и инактивируя продукты, выделяемые базофилами. В эозинофилах содержатся катионные белки, которые активируют компоненты калликреин-кининовой системы и оказывают влияние на свертывание крови. Моноциты циркулируют от 36 до 104 ч, а затем мигрируют в ткани, где образуют обширное семейство тканевых макрофагов. Моноциты и макро- фаги вместе образуют систему мононуклеарных фагоцитов (СМФ). Клетки, объединяемые в фагоцитирующую систему, включают костномозговые предшественники, пул циркулирующих в крови моноцитов и органо- и тканеспецифические макрофаги. Моноциты и макрофаги выполняют многообразные функции: - являются активными фагоцитами, распознают Аг и переводят его в так называемую иммуногенную форму (выполняют функции антигенпрезентирующих клеток), - играют существенную роль в противоинфекционном и противораковом иммунитете; - участвуют в метаболизме липидов и железа; - синтезируют отдельные компоненты системы комплемента и факторы, принимающие участие в сосудисто-тромбоцитарном гемостазе, процессе свертывания крови и растворении кровяного сгустка. Лимфоциты, как и другие виды лейкоцитов, образуются в костном мозге, а затем поступают в циркуляцию. Здесь одна популяция лимфоцитов направляется в вилочковую железу, где в результате контакта со стромальными элементами и гуморальными факторами преимущественно полипептидной природы превращаются в так называемые Т-лимфоциты. У человека окончательное формирование В-лимфоцитов происходит в костном мозге. Популяция Т-лимфоцитов гетерогенна и представлена следующими классами клеток: - T-киллеры, осуществляющие лизис клеток-мишеней, к которым можно отнести возбудителей инфекционных болезней, грибы, микобактерии, опухолевые клетки; - Т-хелперы, или помощники иммунитета. Т-хелперы разделяются на 2 клона (Txl и Tx2). Txl являются регуляторами клеточного, а Tx2 - гуморального иммунитета. Предположение о том, что среди Т-лимфоцитов имеется популяция супрессоров, не нашло экспериментального подтверждения. Большинство В-лимфоцитов в ответ на действие чужеродных антигенов переходят в плазмоциты и продуцируют антитела или иммуноглобулины, т.е. являются антителопродуцентами. Однако среди В-лимфоцитов также различают В-киллеры и В-хелперы. Существует группа клеток, получивших наименование ни Т-, ни В-лимфоциты. К ним относятся так называемые 0-лимфоциты, являющиеся в основном предшественниками Т- и В-клеток и составляющие их резерв. 0-лимфоцитам также относят клетки, носящие название натуральные (природные) киллеры, или НК-лимфоциты. Двойные клетки несут на своей поверхности маркеры и Т-, и В-лимфоцитов и способны заменять как те, так и другие. В норме и патологии учитывается не только количество лейкоцитов, но и их процентное соотношение, получившее наименование лейкоцитарной формулы, или лейкограммы. Лейкоцитарная формула - процентное содержание различных форм лейкоцитов к общему числу лейкоцитов, которое равняется 100%. В крови здорового человека могут встречаться зрелые и юные формы лейкоцитов, однако в норме обнаружить их удается лишь у самой многочисленной группы - нейтрофилов. К ним относятся юные и палочкоядерные нейтрофилы. Увеличение количества юных и палочкоядерных нейтрофилов свидетельствует об омоложении крови и носит название сдвига лейкоцитарной формулы влево; снижение количества этих клеток говорит о старении крови и называется сдвигом лейкоцитарной формулы вправо.  Количество лейкоцитов у новорожденных повышено и равно 10- 30 *109 /л. Число нейтрофилов составляет -60,5 %, эозинофилов – 2%, базофилов -02 %, моноцитов -1,8 %, лимфоцитов – 24 %. В течении первых 2 недель количество лейкоцитов сокращается до 9 – 15 *109 /л, к 4 годам уменьшается до 7-13*109 /л, а к 14 годам достигает уровня, характерного для взрослого. Соотношение нейтрофилов и лимфоцитов меняется, что обуславливает возникновение физиологических перекрестов Первый перекрест. У новорожденного соотношение содержания этих клеток такое же, как и у взрослого. В последующем содержание Нф падает, а Лмф возрастает, так что на 3-4 сутки их количество уравнивается. В дальнейшем количество Нф продолжает снижаться и к 1-2 годам достигает 25 %. В этом же возрасте количество Лмф- 65 %. Второй перекрест. В течении следующих лет число Нф постепенно повышается, а Лмф –снижается, так что у детей в 4 года эти показатели снова уравниваются и составляют по 35 % от общего количества лейкоцитов. Количество Нф продолжает увеличиваться, а количество Лмф – уменьшается, и к 14 годам эти показатели соответствуют таковым у взрослого (4-9 *109 /л). |