Монография исследование водорослей.. Во́доросли. Аутентичного материала
 Скачать 2.99 Mb. Скачать 2.99 Mb.
|

|
600 000 – 750 000 знаков – (250-300 стр.) - аутентичного материала (монография, материалы научных конференций, научные статьи и т.д. не ранее 2014 года) ; Темы связанные с биологией, в частности водорослями и всем что и ними связано Аутентичный(оригинальный английский, учёным из англоговорящей страны) Найти, подготовить и оформить. Клиент: на антиплагиат не проверяется. Весь текст это ведь чужие статьи или книги или ещё что:) Важно чтобы авторы были англоговорящие И общее количество символов или страниц И тематика Ученые впервые обнаружили на территории России штамм зеленой одноклеточной водоросли Coelastrella multistriata. Она представляет большой интерес для биотехнологии, поскольку до 50% ее массы составляют жирные кислоты, которые перспективны в качестве сырья для фармацевтических препаратов, а также производства биодизельного топлива. Результаты исследования, поддержанного грантом Российского научного фонда (РНФ), опубликованы в журнале Scientific Reports. Род Coelastrella объединяет одноклеточных зеленых водорослей, которые обитают на суше и в пресных водоемах Европы. Ученые выяснили, что представители этого рода накапливают в клетках большое количество липидов в виде жирных кислот и каротиноидов (предшественников витамина А) — до 50% и 2% биомассы соответственно. Эти вещества очень ценятся в фармацевтической промышленности, поскольку из них можно синтезировать биологически активные вещества, витамины и пищевые добавки. Исследователи из Института физиологии растений имени К.А. Тимирязева РАН (Москва) с коллегами описали новый штамм водоросли Coelastrella multistriata, а также изучили состав, количество и особенности накопления в ее клетках жирных кислот. Штамм MZ-Ch23 биологи обнаружили в почве на территории Гуровского карьера в Тульской области. Водоросль очистили от сопутствующих микроорганизмов и выращивали в колбах на минеральной среде. Количество и тип жирных кислот, накапливаемых в клетке, исследователи определяли биохимически. Оказалось, что по количеству преобладали ненасыщенные кислоты, в молекулах которых между некоторыми атомами углерода образуются двойные связи. Среди них больше всего оказалось омега-3 и омега-6 жирных кислот, таких как альфа-линоленовая — 38%, гексадекатриеновая — 19% и линолевая — 12%. Омега-3 и омега-6 жирные кислоты очень полезны для человека: на их основе в организме синтезируются многие биологически активные вещества и гормоны, поэтому Coelastrella может стать хорошим источником для их промышленного получения. Также ученые создали для водоросли условия голодания — штамм поместили в среду, в которой не было азота, фосфора или обоих этих элементов. Азот и фосфор необходимы для роста всех микроорганизмов, в том числе одноклеточных водорослей, поскольку с помощью этих элементов собираются «строительные блоки» белков и нуклеиновых кислот. Если же появляется недостаток азота или фосфора, водоросли начинают накапливать запасные питательные вещества, в первую очередь липиды. Поэтому биологи предположили, что при голодании штамм Coelastrella multistriata будет синтезировать еще больше ценных жирных кислот. Оказалось, что в таких условиях водоросли вместо омега-3 и омега-6 стали синтезировать преимущественно насыщенные и мононенасыщенные (только с одной двойной связью) жирные кислоты — до 61% от общего количества. «Наше исследование показало, что общее содержание жирных кислот в клетках Coelastrella multistriata MZ-Ch23 достигает 57% биомассы, что гораздо выше, чем у других видов этого рода водорослей. Поэтому этот штамм перспективен для получения омега-3 и омега-6 жирных кислот, которые в большом количестве накапливаются в клетках при выращивании на полной питательной среде. Кроме того, мы показали, что при голодании водоросль синтезирует насыщенные жирные кислоты, которые можно использовать для производства биодизельного топлива. В дальнейшем мы планируем исследовать влияние других стрессоров, например тяжелых металлов или избыточного освещения, на накопление липидов этим штаммом», — рассказывает руководитель проекта по гранту РНФ Евгений Мальцев, кандидат биологических наук, доцент, ведущий научный сотрудник Лаборатории молекулярной систематики водных растений Института физиологии растений имени К.А. Тимирязева РАН. Во́доросли (лат. Algae) — гетерогенная экологическая группа преимущественно фотоавтотрофных одноклеточных, колониальных или многоклеточных организмов, обитающих, как правило, в водной среде, в систематическом отношении представляющая собой совокупность многих отделов. Вступая в симбиоз с грибами, эти организмы в ходе эволюции образовали совершенно новые организмы — лишайники. Наука о водорослях называется альгологией. Водоросли отличаются большим разнообразием. По разным оценкам, количество видов водорослей составляет от 30 тыс. до более 1 млн [Guiry]. И хотя не определены объемы групп водорослей и часто само понятие вида в различных отделах и классах, по самым консервативным оценкам насчитывается около 72 500 видов водорослей. Традиционно водорослями называют организмы, осуществляющие оксигенный фотосинтез, у которых отсутствуют стерильные ткани вокруг половых репродуктивных структур [Bold, 1973]. Термин «водоросли» является собирательным для огромной группы эукариотических организмов и прокариот, имеющих разное происхождение и не обязательно родственных друг другу. Существует несколько отдельных филогенетических ветвей эукариот, которые эволюционировали независимо друг от друга и содержат фотосинтезирующие организмы. В соответствии с современными представлениями зеленые и харовые водоросли, наряду с настоящими наземными растениями, образуют подцарство Viridiplantae, которое, в свою очередь, вместе с красными водорослями и глаукофитами (вместе образующими подцарство Biliphyta) формирует обособленную ветвь в филогенетическом дереве — царство Archaeplastidae или Plantae [Cavalier-Smith, 1998; Keeling; Lewis, McCourt]. Несмотря на то, что современные генетические данные говорят о том, что водоросли не представляют собой отдельную филетическую группу, а являются эукариотами различного происхождения, получившими в качестве эндосимбионта цианобактерию или образовавшиеся в результате эндосимбиогенеза разных эукариотических организмов, термин «водоросли» является очень удобным, и, в соответствии с классическим пониманием этого термина, мы будем относить к водорослям как эукариотические, так и прока-риотические фотосинтетические организмы, характеризующиеся отсутствием тканей и органов. Общие сведения Водоросли — группа организмов различного происхождения, объединённых следующими признаками: наличие хлорофилла и фотоавтотрофного питания; у многоклеточных — отсутствие чёткой дифференцировки тела (называемого слоевищем, или талломом) на органы; отсутствие ярко выраженной проводящей системы; обитание в водной среде или во влажных условиях (в почве, сырых местах и т. п.). Они сами по себе не имеют органов, тканей и лишены покровной оболочки. Некоторые водоросли способны к гетеротрофии (питанию готовой органикой), как осмотрофной (поверхностью клетки), например жгутиконосцы, так и путём заглатывания через клеточный рот (эвгленовые, динофитовые). Размеры водорослей колеблются от долей микрона (кокколитофориды и некоторые диатомеи) до 30—50 м (бурые водоросли — ламинария, макроцистис, саргассум)[1]. Таллом бывает как одноклеточным, так и многоклеточным. Среди многоклеточных водорослей наряду с крупными есть микроскопические (например, спорофит ламинариевых). Среди одноклеточных есть колониальные формы, когда отдельные клетки тесно связаны между собой (соединены через плазмодесмы или погружены в общую слизь). К водорослям относят различное число (в зависимости от классификации) отделов эукариот, многие из которых не связаны общим происхождением. Также к водорослям часто относят синезелёные водоросли или цианобактерии, являющиеся прокариотами. Традиционно водоросли причисляются к растениям. Водоросли — вездесущие организмы. Они встречаются в раз-личных местообитаниях и играют важную роль в экосистемах. Во-преки бытующему мнению, водоросли распространены не только в воде, но и на суше. Они обычны в поверхностном слое почвы, на поверхности камней, коре деревьев и других увлажненных субстратах. Существуют нивальные виды, встречающиеся в горах на поверхности снега. Цитология Клетки водорослей (за исключением амёбоидного типа) покрыты клеточной стенкой или клеточной оболочкой. Стенка находится снаружи мембраны клетки, обычно содержит структурный компонент (например, целлюлозу) и аморфный матрикс (например, пектиновые или агаровые вещества); также в ней могут быть дополнительные слои (например, спорополлениновый слой у хлореллы). Клеточная оболочка представляет собой или внешний кремнийорганический панцирь (у диатомей и некоторых других охрофитовых), или уплотнённый верхний слой цитоплазмы (плазмалемму), в котором могут быть дополнительные структуры, например, пузырьки, пустые или с целлюлозными пластинками (своеобразный панцирь, тека, у динофлагеллятов). Если клеточная оболочка пластичная, клетка может быть способна к так называемому метаболическому движению — скольжению за счёт небольшого изменения формы тела. Фотосинтезирующие (и «маскирующие» их) пигменты находятся в особых органоидах — хроматофорах. Хроматофор имеет две (красные, зелёные, харовые водоросли), три (эвглены, динофлагелляты) или четыре (охрофитовые водоросли) мембраны. Также он имеет собственный сильно редуцированный генетический аппарат, что позволяет предположить его симбиогенез (происхождение от захваченной прокариотной или, у гетероконтных водорослей, эукариотной клетки). Внутренняя мембрана выпячивается внутрь, образуя складки — тилакоиды, собранные в стопки — граны: монотилакоидные у красных и синезелёных, двух- и больше у зелёных и харовых, трёхтилакоидные у остальных. На тилакоидах, собственно, и расположены пигменты. Хроматофоры у водорослей имеют различную форму (мелкие дисковидные, спиралевидные, чашевидные, звёздчатые и т. д.). У многих в хроматофоре имеются плотные образования — пиреноиды. Это место скопления питательных веществ и зона, в которой наиболее активно идёт процесс фотосинтеза. Продукты фотосинтеза, в данный момент излишние, сохраняются в форме различных запасных веществ: крахмала, гликогена, других полисахаридов, липидов. Помимо прочего липиды, будучи легче воды, позволяют держаться на плаву планктонным диатомовым с их тяжёлым панцирем. В некоторых водорослях образуются газовые пузыри, также обеспечивающие водоросли подъёмную силу. Морфологическая организация таллома У водорослей выделяют несколько основных типов организации таллома: Амёбоидный (ризоподиальный) Одноклеточные организмы, лишённые твёрдой клеточной оболочки, и вследствие этого, не способные сохранять постоянную форму тела. Благодаря отсутствию клеточной стенки и наличию особых внутриклеточных структур клетка способна к ползающему движению посредством псевдоподий или ризоподий. Для некоторых видов характерно образование синцития путём слияния нескольких амёбоидных клеток. Амёбоидное строение могут вторично приобретать некоторые монадные формы путём отбрасывания или втягивания жгутиков. Монадный Одноклеточные водоросли, имеющие постоянную форму тела, жгутик(и), часто стигму, а пресноводные — сократительную вакуоль. Клетки активно двигаются в вегетативном состоянии. Часто встречается объединение нескольких монадных клеток в колонию, окружённую общей слизью, в некоторых случаях они даже соединяются между собой посредством плазмодесм. У высокоорганизованных форм с многоклеточным талломом часто имеются расселительные стадии — зооспоры и гаметы, имеющие монадную структуру. Коккоидный Одноклеточные, лишённые каких-либо органоидов передвижения и сохраняющие постоянную форму тела в вегетативном состоянии клетки. Чаще всего имеется утолщённая клеточная стенка или панцирь, могут быть различные выросты, поры и пр. для облегчения парения в толще воды. Многим водорослям с данной структурой свойственно образование колоний. Некоторые диатомеи и десмидиевые способны к активному передвижению путём выделения слизи. Пальмеллоидный (капсальный) Постоянное, достаточно крупное, как правило, прикреплённое к субстрату, образование из нескольких коккоидных клеток, погружённых в общую слизистую массу. Клетки непосредственно между собой не объединяются — отсутствуют плазмодесмы. Временную стадию жизненного цикла с аналогичной морфологией называют пальмеллевидным состоянием. В такое состояние могут переходить многие монадные и коккоидные водоросли при наступлении неблагоприятных условий, образующиеся при этом пальмеллевидные образования, как правило, мелкие и не имеют постоянной формы. Нитчатый (трихальный) Клетки соединены в нить, простую или разветвлённую. Нити могут свободно плавать в толще воды, прикрепляться к субстрату, либо объединяться в колонию. Вегетативно нитчатые водоросли размножаются обычно распадом нити на отдельные фрагменты. Рост нитей может идти четырьмя путями: диффузным — делятся все клетки нити, интеркалярным — зона роста расположена в середине нити, апикальным — делением конечных клеток, и базальным — делением клеток у основания таллома. Клетки в нити не имеют жгутиков и могут быть связаны между собой плазмодесмами. Разнонитчатый (гетеротрихальный) Есть две системы нитей: стелющиеся по субстрату горизонтальные и отходящие от них вертикальные. Горизонтальные нити тесно смыкаются, либо могут сливаться в псевдопаренхиматозную пластинку и выполняют, в основном, опорную функцию и функцию вегетативного размножения, вертикальные нити — преимущественно ассимиляторную функцию. Иногда может наблюдаться редукция, либо чрезмерное развитие тех или иных нитей, приводящее к вторичной утрате или нарушению характерных черт гетеротрихального строения (при редукции вертикальных нитей, например, таллом может представлять собой простую однослойную пластинку, полностью прикреплённую к субстрату). Пластинчатый Многоклеточные талломы в форме пластинок из одного, двух или нескольких слоёв клеток. Возникают при продольном делении клеток, составляющих нить. Число слоёв зависит от характера образования перегородок при делении клеток. Иногда слои могут расходиться, и таллом тогда приобретает трубчатую форму (полый внутри), стенки при этом становятся однослойными. Сифональный (неклеточный, сифоновый) Отсутствуют клеточные перегородки, в результате чего таллом, часто крупный и внешне дифференцированный, формально представляет собой одну клетку с большим количеством ядер. Сифонокладальный Таллом представлен многоядерными клетками, соединёнными в нитчатые или иной формы многоклеточные талломы (Siphonocladales). Харофитный (членисто-мутовчатый) Свойственна только харовым водорослям. Таллом крупный, многоклеточный, состоит из главного побега с ветвями и отходящими от него, иногда ветвящимися, членистыми боковыми побегами. Боковые побеги отходят от главного в области узлов, часть побега между узлами состоит, как правило, из одной крупной клетки и называется междоузлием. Сарциноидный Колонии, представляющие собой группы (пачки или нитеобразные образования), которые возникают в результате деления одной исходной клетки и заключены в растягивающуюся оболочку этой клетки. Псевдопаренхиматозный (ложнотканевый) Представлен слоевищами, которые образовались в результате срастания разветвлённых нитей, нередко сопровождаемого морфофункциональной дифференциацией получающихся ложных тканей. У части синезелёных, зелёных и красных водорослей в слоевище откладываются соединения кальция, и оно становится твёрдым. Водоросли лишены корней и поглощают нужные им вещества из воды всей поверхностью. Крупные донные водоросли имеют органы прикрепления — подошву (уплощённое расширение в основании) или ризоиды (разветвлённые выросты). У некоторых водорослей побеги стелются по дну и дают новые слоевища.  Рис. 4. Морфологическая организация талломов водорослей: а — монадный (Chlamydomonas); б — амебоидный (Chrysamoeba); в — пальмеллоид-ный (Hydrurus); г — коккоидный (Pediastrum); д — сарциноидный (Chlorosarcina); е — нитчатый (Ulothrix); ж — разнонитчатый (Fritschiella); з, и — тканевой (Porphyra, Laminaria); к — сифональный (Caulerpa); л — сифонокладальный (Cladophora) [по: Арбузова, Левенец] Строение клеток водорослей. Клетки одноклеточных и многоклеточных водорослей имеют ряд морфолого-анатомических особенностей, определяющих их функциональное предназначение. Наружные покровы. Наружные покровы клеток водорослей характеризуются значительным разнообразием. Существуют группы водорослей, протопласт которых окружен только плазмалеммой. Такие покровы характерны для одноклеточных водорослей, имеющих амебоидную жизненную форму, а также гамет и зооспор многоклеточных водорослей. У большинства водорослей клетки покрыты клеточной оболочкой (стенкой). Химический состав и строение клеточных оболочек зависят от систематической принадлежности водоросли. Клетки цианобактерий покрыты четырехслойной клеточной стенкой, ос-новным компонентом которой является пептидогликан — муреин, не встречающийся у эукариотических организмов. Клеточная стенка зеленых водорослей образована целлюлозой, в которой может откладываться известь (рис. 5). Целлюлозная оболочка характерна также для динофитовых водорослей, но, в отличие от зеленых, их оболочка образована системой пластинок, располагающихся внутри клеток в пузырьках — везикулах. У золотистых, желтозеленых и диатомовых водорослей оболочка клетки окремневает. Клет-ки эвгленовых и криптомонад окружены белковой пелликулой. 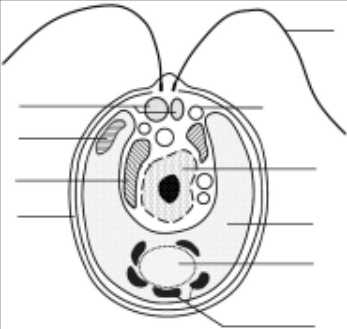 Рис. 5. Строение Chlamydomonas sp.: 1 — сократительная вакуоль; 2 — глазок; 3 — митохондрия; 4 — клеточная стенка; 5 — вакуоль; 6 — жгутики; 7 — ядро; 8 — хлоропласт; 9 — пиреноид; 10 — крах-мальные зерна У прокариотических клеток отсутствуют ограниченные мем-браной органеллы (ядро, пластиды, митохондрии, диктиосомы, тельца Гольджи, жгутики), что характерно для цианобактерий. Мезокариотическая клетка (панцирные жгутиконосцы из группы динофлагеллят) несет четко ограниченное ядро, но в его строении сохраняются примитивные черты. Эукариотические водоросли обычно содержат одно четко дифференцированное ядро, хотя существуют виды, у которых количество ядер увеличивается. Так, для кладофоры характерны клетки с несколькими десятками ядер, еще больше ядер у водяной сеточки (сотни ядер). Ядро содержит генетический материал клетки, ядрышко, кариоплазму и окружено двойной мембраной с порами в ней. Митохондрии водорослей имеют овальную форму и отделены от цитоплазмы двухслойной оболочкой. Внутренняя оболочка формирует большое количество выростов в виде крист. Хлоропласты водорослей крайне разнообразны по форме, могут быть кольцевидными (Ulotrix), чашевидными (Chlamydomonas), цилиндрическими, спиральными (Spirogyra), пластинчатыми (некоторые диатомовые), дисковидными или округлыми (некоторые зеленые водоросли). Располагаются обычно вдоль стенок, но также могут заполнять цитоплазму (зеленые, бурые, красные водоросли) или локализоваться в центре клетки (Zygnema, десмидиевые). Хлоропласты водорослей также отличаются по числу оболочек, характеристике и расположению тилакоидов, составу хлорофиллов и пигментов (табл. 1). Хлоропласт может иметь две (красные, зеленые, харовые водоросли), три (эвгленовые и динофлагелляты) или четыре (охрофиты, криптофиты и хлорарахниофиты) внешние мембраны. Внутренняя мембрана хлоропластов имеет многочисленные выпячивания, называемые тилакоидами, собранные в граны, в которых осуществляется фотосинтез. Количество тилакоидов в гранах варьирует и зависит от систематической принадлежности водоросли (табл. 1). Большинство хлоропластов зеленых водорослей несут пиреноиды — тельца, содержащие большое количество рубискокарбоксилазы (обычно говорят или рубиско- или рибулозобифосфаткарбоксилаза), ответственные за концентрацию углерода [Villareal, Renner], вокруг которых откладывается крахмал. Таблица 1 |