Шпора к экзамен норфиз. шпора к экзам по норм физе. Билет 1 Нервная регуляция висцеральных функций эффекты с висцерорецепторов, классификация висцерорецепторов, висцерорефлексов, уровни замыкания рефлекторных дуг.
 Скачать 1.45 Mb. Скачать 1.45 Mb.
|

|
Состав и свойства желудочного сока. Клетки: главные – вырабатывают ферменты; париетальные – HCl, добавочные – муцин. В сутки выделяется 1,5 – 2,0л. желудочного сока. Удельный вес 1002 – 1007, рН – 0,8 – 1,5, HCl содержится 0,3 – 0,5%, Н2О – 99,0 – 99,5%, 1,0 – 0,5% плотных органических и неорганических веществ (хлориды, сульфаты, фосфаты, бикарбонаты Na, К, Са, Mg). В небольшом количестве содержится: мочевина, мочевая кислота и др. Секреция зависит от количества и состава пищи: количество сока убывает: мясо → хлеб → молоко кислотность снижается: белки → углеводы → жиры. Фазы желудочной секреции. 1) Сложнорефлекторная: - условнорефлекторная – до приема пищи при раздражении органов чувств (вид, запах); - рефлекторная – пища во рту, раздражение рецепторов ротовой полости → активируется, н.V → увеличивается секреция. Сока выделяется много. Это аппетитный сок. 2) Желудочная фаза. Пища в желудке. Различают: нервную регуляцию → пища действует на механорецепторы → н.V → повышение секреции; гуморальную – это экстрактивные вещества из мяса, овощей → железы → ↑ секреция, бомбезин, гистамин. Действие гастрина → увеличивает образование HCl. Образуется из прогастрина под действием АХ и продуктов гидролиза белка. 3) Кишечная фаза. Нервная регуляция – поступление в кишечник недостаточно обработанной пищи → механорецепторы → н. V → усиление секреции в желудке. Гуморальная регуляция – энтерогастрин → усиливает секрецию в желудке. Экстрактивные вещества, образующиеся при пищеварении в 12п. кишке активизируют секрецию в желудке. Торможение секреции в желудке: а) рефлекторным путем: - с хеморецепторов и механорецепторов 12 перстной кишки – энтерогастральный рефлекс, эмоции тормозят секрецию. б) гуморальным путем – тормозят секрецию: продукты гидролиза жира, полипептиды, АК, холецистокинин, секретин. 3. Транспорт газов кровью. Содержание кислорода и углекислоты в крови. Транспортные формы О2 иСО2. Факторы, влияющие на транспорт газов кровью. Кривая диссоциации гемоглобина. Перенос кислорода: 1) Физически растворенный 0,3мл в 100мл плазмы; 2) Растворенный О2 диффундирует в эритроциты и соединяется с Нв, образуется НвО2 – оксигемоглобин. В таком виде кривой его соединяется 18 – 20 об% или 180 – 200 мл в 1000 мл крови. 3) КСК = Нв ∙ 1,34 мл. Факторы влияющие на образование НвО2. 1) Напряжение О2 в крови. Графически эту зависимость можно представить в виде кривой диссоциации оксигемоглобина. 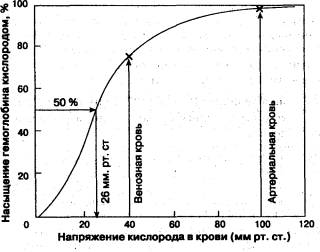 При напряжении О2 = 0 Нв = 0. Повышение содержания О2 вызывает не совсем пропорциональный рост количества НвО2. Она носит S – образный характер. При напряжении О2 = 0 Нв = 0. Повышение содержания О2 вызывает не совсем пропорциональный рост количества НвО2. Она носит S – образный характер.Количество НвО2 быстро нарастает до 80% при повышении рО2 с 10 до 40мм рт ст. При 60мм рт ст. Нв насыщается О2 на 90%. При дальнейшем увеличении рО2 количество НвО2 увеличивается до 96%. Фактически кривая диссоциации оксигемоглобина показывает сродство Нв к О2 в зависимости от различных факторов: 1) Снижение сродства Нв к О2вызывает снижение рН (закисление крови). Кривая диссоциации НвО2 сдвигается вправо. 2) Увеличение СО2. в митохондриях – снижение сродства (эффект Вериго). 3) Повышение tоснижает сродство. 4) Повышение активности 2 – 3 дифосфоглицерата (фермент в эритроците, усиливающий отдачу О2 гемоглобином – гипоксия). При работе тканей все эти факторы вызывают распад НвО2. Транспортные формы СО2. 1) в виде Н2СО3 – 25мл СО2 + Н2О ← → Н2СО3 ↓ карбангидраза 2) в виде карбгемоглобина – 50мл. 3) в виде Na соли угольной кислоты в плазме и К соли в эритроцитах – 480мл. 4) в растворенном в плазме виде.25/580мл.
4. Биле т №7 1. Рефлекторный принцип деятельности ЦНС. Рефлекторная дуга. Рецептивное поле рефлекса. Примеры рефлексов. Основное положение рефлекторной теории заключается в утверждении, что деятельность организма есть закономерная рефлекторная реакция на стимул. Рефлекс реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием центральной нервной системы. В естественных условиях рефлекторная реакция происходит при пороговом, надпороговом раздражении входа рефлекторной дуги — рецептивного поля данного рефлекса. Рецептивным полем называется определенный участок воспринимающей чувствительной поверхности организма с расположенными здесь рецепторными клетками, раздражение которых инициирует, запускает рефлекторную реакцию. Рецептивные поля разных рефлексов имеют определенную локализацию, рецепторные клетки — соответствующую специализацию для оптимального восприятия адекватных раздражителей (например, фоторецепторы располагаются в сетчатке; волосковые слуховые рецепторы — в спиральном (кортиевом) органе; проприорецепторы — в мышцах, в сухожилиях, в суставных полостях Структурной основой рефлекса является рефлекторная дуга — последовательно соединенная цепочка нервных клеток, обеспечивающая осуществление реакции, или ответа, на раздражение. Рефлекторная дуга состоит из афферентного, центрального и эфферентного звеньев, связанных между собой синаптическими соединениями. В зависимости от сложности структуры рефлекторной дуги различают моно- и полисинаптические рефлексы. Еще одним звеном рефлекса является — петля обратной связи, установливающая связь между реализованным результатом рефлекторной реакции и нервным центром, выдающим исполнительные команды. Классификации рефлексов. Существуют различные классификации рефлексов: По способу вызывания различают безусловные рефлексы (категория рефлекторных реакций, передаваемых по наследству) и условные рефлексы (рефлекторные реакции, приобретаемые на протяжении индивидуальной жизни организма). Различают экстероцептивные рефлексы — рефлекторные реакции, инициируемые раздражением многочисленных экстерорецепторов (болевые, температурные, тактильные и т. д.), интероцептивные рефлексы (рефлекторные реакции, запускаемые раздражением интероцепторов: хемо-, баро-, осморецепторов и т. д.), проприоцептивные рефлексы (рефлекторные реакции, осуществляемые в ответ на раздражение проприорецепторов мышц, сухожилий, суставных поверхностей и т. д.). В зависимости от уровня активации части мозга дифференцируют спинномозговые, бульбарные, мезенцефальные, диэнцефальные, кортикальные рефлекторные реакции. По биологическому назначению рефлексы делят на пищевые, оборонительные, половые и т. д. Классфикация Когана: Элементарные безусловные рефлексы, представлены простыми рефлекторными реакциями, осуществляемыми на уровне отдельных сегментов спинного мозга. Они имеют местное значение, вызываются локальным раздражением рецепторов данного сегмента тела и проявляются в виде локальных сегментарных сокращений поперечнополосатой мускулатуры. Роль: обеспечении простейших приспособительных реакций к внешним воздействиям местного значения, а также в приспособительных изменениях отдельных внутренних органов. Координационные безусловные рефлексы согласованные акты локомоторной деятельности или комплексные реакции вегетативных функциональных объединений внутренних органов, вызываются раздражением определенных групп внешних или внутренних рецепторов, однако их эффект не ограничивается локальной реакцией путем последующей активации широкого класса экстеро-, интеро- и проприорецепторов, а формирует сложные координационные акты сокращения и расслабления, возбуждения или торможения деятельности ряда внутренних органов. Интегративные безусловные рефлексы. Вызываются например пищевыми, болевыми раздражителями. Пример такой реакции — ориентировочная реакция. Биологическое значение которой заключается в перестройке организма, которая обеспечивает оптимальную подготовку к восприятию и быстрому анализу нового неизвестного сигнала в целях организации рационального ответа.. Сложнейшие безусловные рефлексы (инстинкты) представляют собой видовые стереотипы поведения, организующиеся на базе интегративных рефлексов по генетически заданной программе. Сложные формы высшей нервной деятельности представлены психическими реакциями, в качестве вызывающих подобные реакции стимулов обычно выступают сложные комплексные раздражители. Часто такие рефлекторные реакции имеют усеченную рефлекторную дугу (отсутствует эфферентное звено рефлекторной дуги). 2. Виды сокращений желудка. Регуляция сокращений желудка. Сокращения бывают 1) тонические а) в пустом б) в наполненном желудке 2) периодические а) в пустом желудке → натощак, голодные сокращения б) наполненный → обычной пищей и грубой пищей Характеристика сокращений. Тонические: а) пустой желудок – пластический тонус (50мл.3). б) наполненный – объемная адаптация (до 3л.). Периодические сокращения: пустой желудок. 1) натощак – 5 – 6 сокращений в минуту, давление 8 мм. рт. ст. 2) голодные сокращения – в пилорической части через 1,5 – 2 часа, продолжительность 15 – 20 минут, амплитуда 80 – 100 мм. рт. ст. Наполненный желудок. 2 типа сокращений. I тип – после приема пищи частота – 5 – 6 в минуту. амплитуда – 5 – 8 в минуту. II тип – через 1,5 – 2 часа после приема пищи. Обеспечивает эвакуацию химуса из желудка. Волна начинается в кардиальной части и распространяется до пилорического, не затухая. Давление 80 – 100, продолжительность волны 10 – 30 секунд. Регуляция сокращений желудка. 1) Рефлекторная: а) Безусловнорефлекторная. Рецепторное поле в полости рта, пищевода, желудка, двенадцатиперстной кишке, толстой кишке. Дуга может замыкаться на различных уровнях 1. МСНС (местный уровень регуляции). 2) вегетативные ганглии; 3) на уровне нервных центров симпатической и парасимпатической систем. Эфферентное влияние: n. V. – усиливает; симпатическая система вызывает снижение моторики и угнетение эвакуации из желудка. Условнорефлекторные механизмы ( на вид, запах, разговор пище). Гуморальная регуляция – гастроинтестинальные гормоны. Усиление моторики – мотилин, гистамин, серотонин Торможение – катехоламины, холецистокинин, соматостатин. 3. Дыхательный центр (понятие). Отделы ДЦ и их функции (бульбо-понтийный, спинномозговой, гипоталамический, корковый отделы). 1) Дыхательный центр. Это совокупность нейронов, обеспечивающих координацию деятельности дыхательной мускулатуры и приспособление деятельности дыхательной системы к изменившимся условиям. По последним представлениям о нервных центрах. ДЦ располагается на различных уровнях ЦНС: спинном, БПО, ЛРК, кора. Роль различных отделов в регуляции дыхания. I) Ствол мозга – здесь находится жизненно важный отдел ДЦ – бульбо – понтийный. Варолиев мост – в передней части находятся нейроны, обладающие тонической активностью, называется пневмотаксический центр. Роль: 1.обеспечивает смену дыхательных фаз (вдох – выдох); 2. увеличивает скорость развития вдоха; 3. повышает возбудимость нейронов выключающих вдох (нарушение связи). II)Продолговатый мозг. Инспираторные нейроны образуют центр вдоха. В нем можно выделить 2 отдела (воспринимающий и эффекторный). Функции инспираторных нейронов: 1) самовозбуждаются, но при условии: а) их связи с другими нейронами ДЦ, среди которых, возможно, есть пейсмекеры; б) наличии афферентных сигналов или сигналов с хеморецепторов. 2) воспринимают сигналы от хеморецепторв; 3)передают сигналы к инспираторным мышцам. Экспираторные нейроны образует центр выдоха преимущественно в вентральных ядрах. Также 2 части: воспринимающая и исполнительная. Функции экспираторных нейронов. 1) воспринимают сигналы от механорецепторов легких. 2) от проприорецепторов дыхательных мышц. 3) тормозят инспираторные нейроны, обеспечивая смену вдоха на выдох. III)Гипоталамус: 1) автоматизированное управление через АНС и ЖВС при поступлении сигналов - с интерорецепторов; - с проприорецепторов; - с терморецепторов (тепловая одышка, растет ЧД и отдача тепла). Лимбическая система изменяет дыхание при поведенческих реакциях. Кора БП: 1) тормозит ДИ; 2) условные рефлексы; 3) произвольная регуляция. 4. Билет №8 1. 2. Переход содержимого желудка в кишечник. Пищеварение в 12ти перстной кишке. Переход содержимого желудка в двенадцатиперстную кишку, или эвакуация, вызывается чередующимися открытием и закрытием пилорического сфинктера. Механизм - когда содержимое желудка, пропитанное желудочным соком, поступает в его пилорическую часть, рецепторы в этом месте раздражаются HCl и сфинктер открывается. Часть содержимого желудка переходит в двенадцатиперстную кишку. Реакция в кишечнике становится кислой вместо щелочной, и теперь та же соляная кислота, действуя на рецепторы слизистой оболочки двенадцати перстной кишки, рефлекторно вызывает закрытие пилорического сфинктера. Когда под влиянием щелочных соков (поджелудочный и кишечный соки, желчь) соляная кислота нейтрализуется, а большая часть поступившей массы переместится дальше по кишечнику, весь процесс повторяется снова. Одна из основных функций двенадцатиперстной кишки заключается в приведении pH поступающей из желудка пищевой кашицы к щелочному, не раздражающему более дистальные отделы тонкой кишки и пригодному для осуществления кишечного пищеварения. Именно в двенадцатиперстной кишке и начинается процесс кишечного пищеварения. Другая важнейшая функция двенадцатиперстной кишки состоит в инициации и регулировании секреции панкреатических ферментов и жёлчи в зависимости от кислотности и химического состава поступающей в неё пищевой кашицы. Третья важнейшая функция двенадцатиперстной кишки заключается в поддержании обратной связи с желудком — осуществлении рефлекторного открывания и закрывания привратника желудка в зависимости от кислотности и химизма поступающей пищевой кашицы, а также регулировании кислотности и пептической активности секретируемого в желудке сока через секрецию гуморальных факторов, влияющих на секреторную функцию желудка. 3. Современные представления о механизмах периодичности дыхания (генератора центрального инспираторного возбуждения, механизм выключения инспирации, пневмотаксический центр, механорецепторы легких) Главной особенностью дыхательного центра является периодичность, при которой возбуждение нейронов изменяется их торможением. В основе периодичности лежит функция бульбарного отдела. При этом решающая роль принадлежит нейронам дорсального ядра. Считают, что они являются своеобразным «водителем ритма». С бульбарного центра поступает возбуждение от многих образований ЦНС, в том числе от пневмотаксичного центра. Так, если перерезать ствол мозга, отделив варолиев мост от продолговатого мозга, то у животных снижается частота дыхательных движений.. Пневмотаксичний и бульбарный центры имеют двусторонние связи, с помощью которых пневмотаксичний центр ускоряет возникновение следующих инспираций и експираций. На активность нейронов дыхательного центра влияют другие отделы ЦНС, такие, как ретикулярная формация, гипоталамус, кора больших полушарий. Например, характер дыхания меняется при эмоциях. Скелетные мышцы, которые участвуют в дыхании, часто выполняют и другие движения. Да и собственное дыхание, его глубину и частоту человек может менять сознательно, что свидетельствует о влиянии на дыхательный центр коры больших полушарий. Благодаря этим связям дыхания сочетается с выполнением рабочих движений, речевой функцией человека. Таким образом инспираторных нейронов, как «водители ритма», существенно отличаются от настоящих пейсмекерных клеток. При возникновении ритмики основных дыхательных нейронов дорсального ядра необходимо учитывать два условия: а) «последовательность поступления» каждой группы нейронов именно этого отдела б) обязательное импульсацию от других отделов ЦНС и импульсацию от различных рецепторов. Дыхание является вегетативной функцией, а выполняется скелетными мышцами. Поэтому механизмы его регуляции имеют общие черты с механизмами регулирования деятельности как вегетативных органов, так и скелетных мышц. Потребность в постоянном дыхании обеспечивается автоматически за счет активности дыхательного центра. Однако вследствие того, что дыхание осуществляют скелетные мышцы, возможны и произвольные изменения характера дыхания, обусловленные влиянием коры больших полушарий на дыхательный центр. Если во внутренних органах (сердце, кишки) автоматизм обусловлен лишь свойствами водителей ритма, то в дыхательном центре периодическая деятельность управляется сложными механизмами. Периодичность обусловлена: 1) согласованной активностью различных отделов дыхательного центра, 2) поступлением сюда импульсов от рецепторов, 3) поступлением сигналов от других отделов ЦНС, в том числе и от коры головного мозга. Кроме того, при анализе механизма периодичности дыхания нужно учесть, что спокойное и форсированное дыхание существенно отличаются по количеству мышц, участвующих в этом акте. Во многом эта разница определяется уровнем привлечения вентрального отдела бульбарного дыхательного центра, в котором есть как инспираторная, так и экспираторная нейроны. При спокойном дыхании эти нейроны относительно малоактивны, а при глубоком дыхании их роль резко возрастает. 4. Билет №9 1.Уровень поляризации клеточных мембран как регулятор функционального состояния ткани. Зависимость возбудимости от потенциала покоя. Изменение возбудимости при потенциале действия. |