биохимия хазипов. Биохимия животных Электронный дидактический комплекс (эдк)
 Скачать 14.38 Mb. Скачать 14.38 Mb.
|

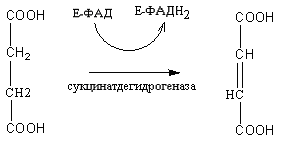
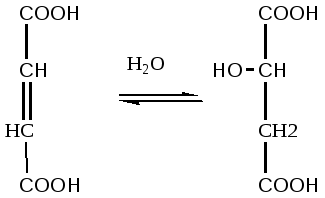
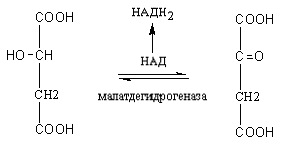
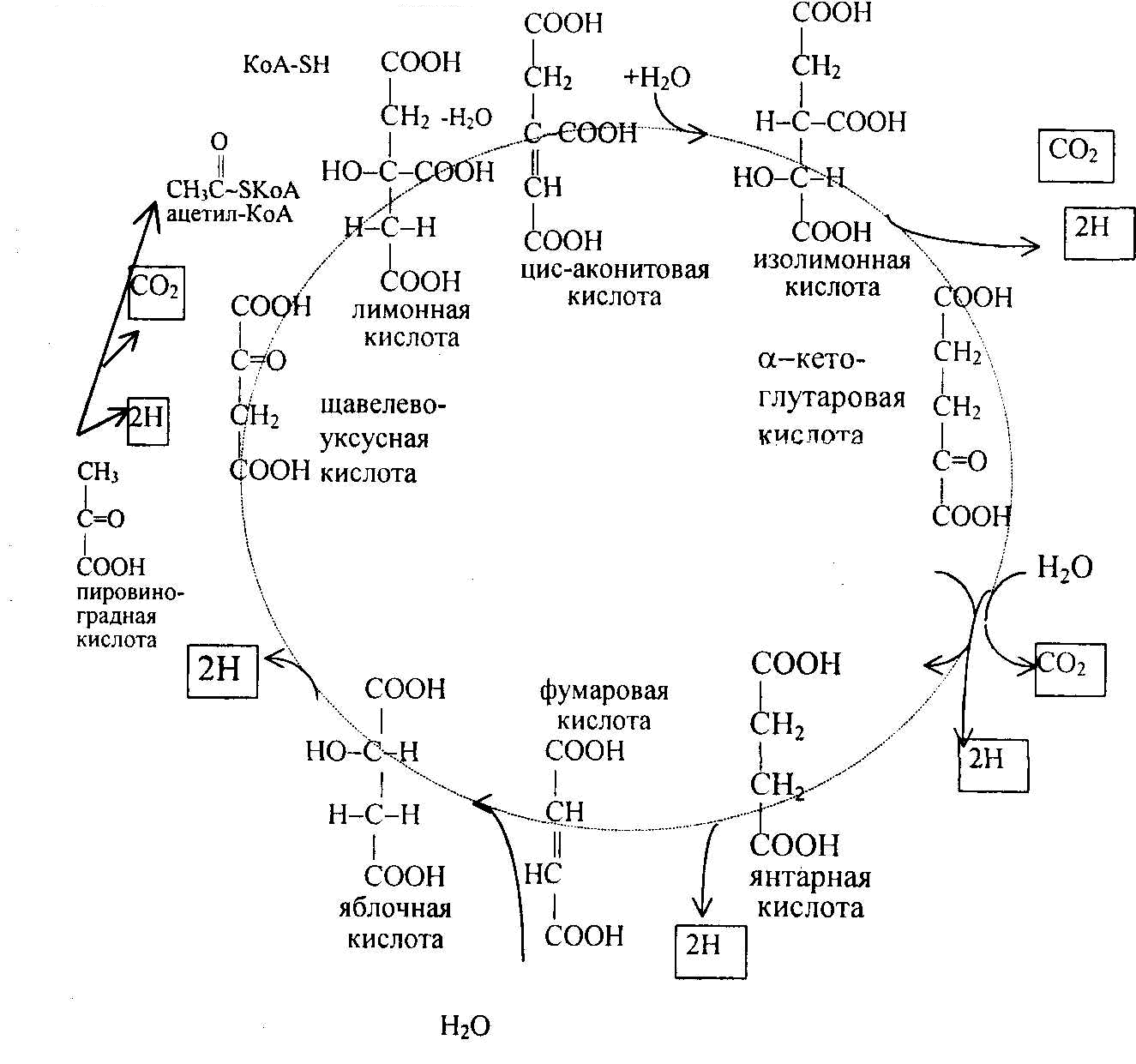
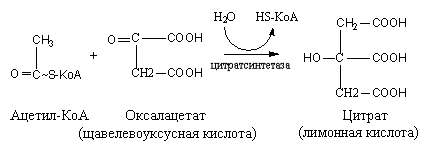
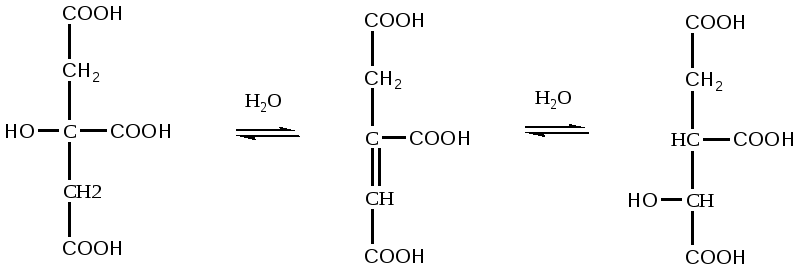
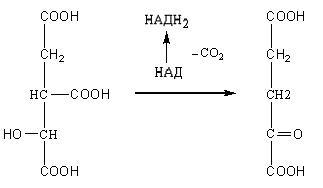
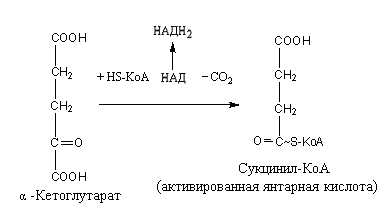
КоА до конечных продуктов – CO2, H2O, с выделением энергии. В цикле трикарбоновых кислот происходит Дегидрирование ди- и трикарбоновых кислот, тканевое дыхание и окислительное фосфорилирование, в этих реакциях участвуют ферменты дыхательной цепи и окислительного фосфорилирования.
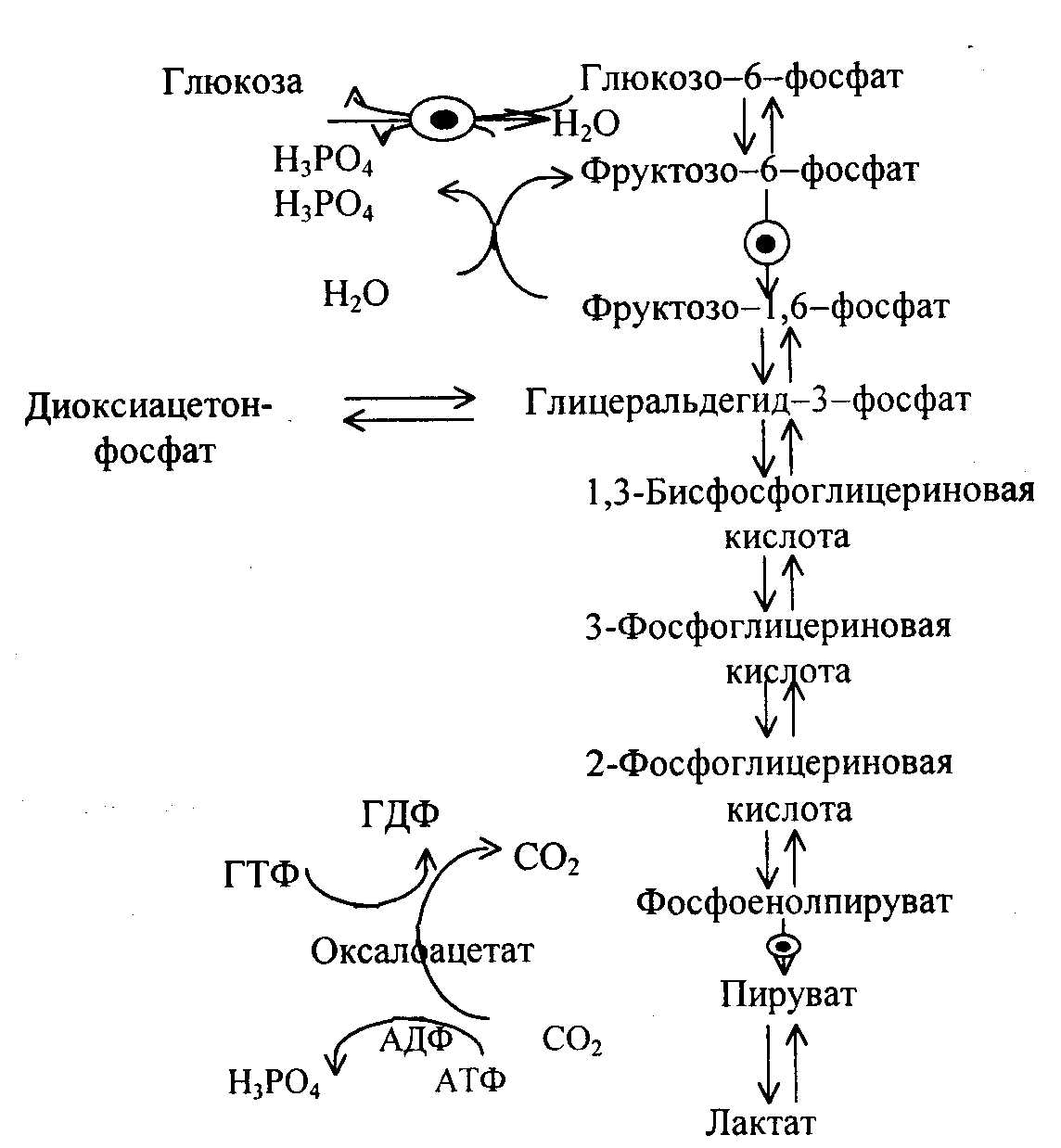
SКоА, 1 молекула углекислоты, 1 молекула воды и 3 молекулы АТФ:8. ХИМИЯ И ОБМЕН УГЛЕВОДОВУглеводы – органические соединения, состоящие чаще всего из углерода, водорода, кислорода. Известно много углеводов, содержащих, кроме того, фосфор, серу, азот. Углеводы широко распространены в природе. Они образуются в растениях в результате фотосинтеза и составляют 80-90 % сухой массы растений. В организме животных содержание углеводов составляет 1-2 % сухого веса ткани. Значение углеводов в организме многогранно. Они являются: 1)основной структурой растительной клетки; 2) источником энергии; 3) откладываются в виде запасных питательных веществ (крахмал). В организме животных и человека углеводы являются источником химической энергии. Отдельные органы удовлетворяют свои потребности в энергии в основном в результате расщепления глюкозы: головной мозг на 80%, сердце на 70-75%. Углеводы откладываются в тканях животного организма в виде запасных питательных веществ (гликоген). Некоторые из них выполняют опорные функции (гиалуроновая кислота), участвуют в защитных функциях, задерживают развитие микробов (мукополисахариды), являются химической основой для построения молекул биополимеров – нуклеиновых кислот, сложных белков, служат составной частью макроэргических соединений. По химическим свойствам углеводы являются полиоксиальдегидами и полиоксикето-спиртами. Различают углеводы простые (моносахариды) и сложные (полисахариды). Моносахариды Моносахариды классифицируют: 1) по наличию альдегидной или кетонной группы (альдозы и кетозы); 2) по числу углеродных атомов - триозы, тетрозы, пентозы, гексозы; 3) по химической природе: нейтральные сахара, кислые сахара, аминосахара. Моносахариды – белые кристаллические вещества, хорошо растворимые в воде, сладкие на вкус, оптически активны. Вступают в химические реакции, характерные для альдегидо- и кетоспиртов:
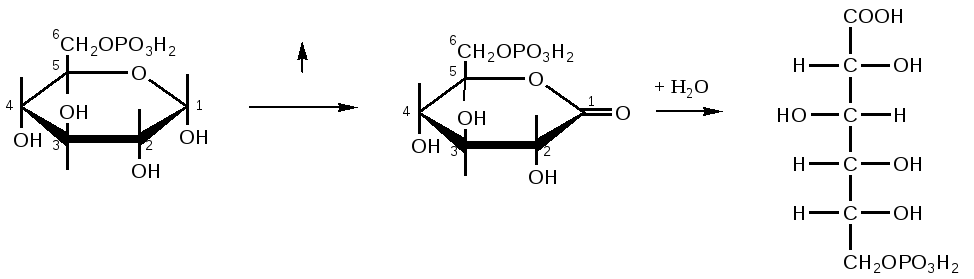
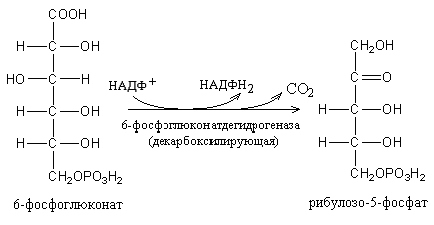
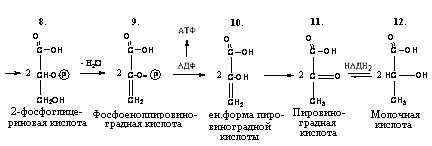
Триозы. Общая формула C3H6O3. По химическим свойствам являются альдегидо- и кетоспиртами. Содержатся в тканях в виде сложных эфиров с ортофосфорной кислотой. Тетрозы – С4H8O4 по химическим свойствам альдозы и кетозы. Наибольшее значение имеет эритроза - продукт пентозного пути окисления углеводов. Пентозы – С5Н10О5, в природе широко распространены, входят в состав сложных соединений, могут существовать в альдо- и кетоформах и в циклической фуранозной форме. Наиболее широко распространена D-рибоза. Фосфорилированная рибоза входит в состав нуклеотидов – составных частей РНК. В тканях находится также кеторибоза D-рибулоза, в виде D-рибулозо-5-фосфата: Кроме того, широко распространена в природе 2-D-дезоксирибоза, входящая в состав ДНК. Ксилоза, ксилулоза, арабиноза имеют значительное распространение. D-арабиноза входит в состав возбудителя туберкулеза. Гексозы – C6H12O6 (рис.8.1) – наиболее широко распространены в природе: составные части моно- и полисахаридов: 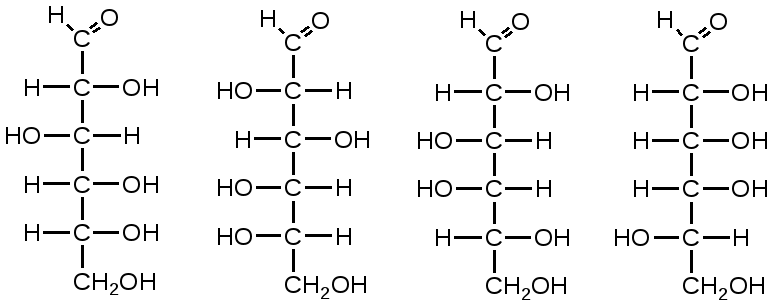 D (+)-глюкоза L(-)-глюкоза D(+)-галактоза L(-)-галактоза 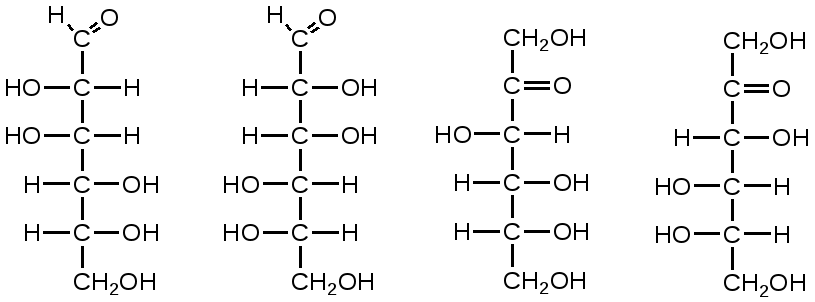 D (+)-манноза L(-)-манноза D(-)-фруктоза L(+)-фруктоза Рис. 8.1. Структурные формулы гексоз. 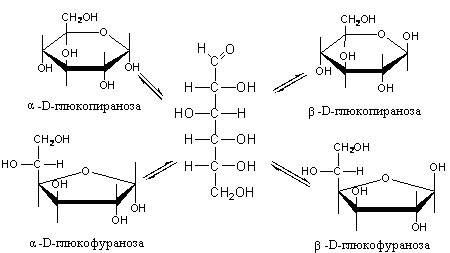 Рис.8.1. Структурные формулы глюкозы D-глюкоза (декстроза, виноградный сахар, виноградный сахар содержит до 20%) – альдогексоза, наиболее часто встречающаяся в пиранозной форме. Они являются структурными компонентами олиго- и полисахаридов в тканях. В крови D-глюкопиранозы содержится 0,04-0,1 %. D-манноза в виде (β-D-маннопиранозы входит в состав белков, гликолипидов. Манноза – составная часть полисахаридов бактерий, дрожжей, гликопротеидов крови. Используется в качестве питательной среды в бактериологии. D-галактоза встречается в виде β-D-галактопиранозы, которая входит в состав молочного сахара (лактозы). Галактоза находится в составе полисахаридов, гликозидов, цереброзидов и т.д. Кроме того, встречается L-лактоза, которая входит в состав агар-агара, полисахаридов, слизи льняного семени. D-фруктоза (левулеза, фруктовый сахар) встречается в свободном виде – кетогексоза во фруктах, меде, овощах и т.д. Фруктоза – самый сладкий сахар, входит в состав сахарозы. Гептозы – С7Н14О7 могут быть в виде альдоз и кетоз, входят в состав полисахаридов. Наиболее распространенной является D-седогептулоза, которая участвует в процессах пентозофосфатного пути окисления углеводов в клетке в виде фосфорных эфиров. 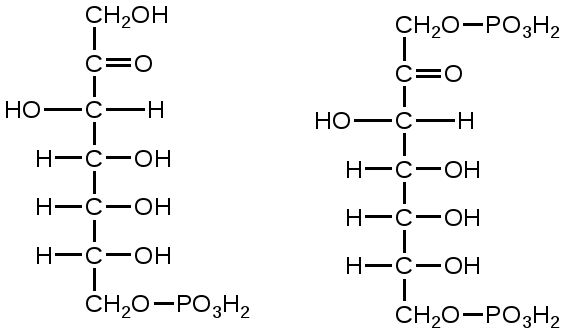 D-седогептулоза-7-фосфат D-седогептулоза-1,7-дифосфат Производные моносахаридов. Уроновые кислоты – производные альдоз, у которых первичная спиртовая группа окислена до карбоксильной. Входят в состав полисахаридов гликозидов. Широко распространена β-D-глюкуроновая кислота (рис.8.2.). Аминосахариды – производные моносахаридов, у которых - ОН группа у С-2 замещена аминогруппой –NH2. Представители – глюкозамин (2-амино-Д-глюкоза), галактозамин (2-амино- D-галактоза). Гексозамины входят в состав ряда белков, веществ группы крови и бактериальных мукополисахаридов. Гликозиды – это эфирообразные соединения, получающиеся в результате замещения радикалом атома водорода в полуацетальном гидроксиле циклической формы моносахарида. Например, метанол в присутствии HCl приводит к образованию α-и β-метил-D-гликозида. Неуглеводная часть молекулы гликозидов, группа, замещающая водород -ОН группы C-1 называется аглюконом. Олигосахариды в природе представлены дисахаридами и трисахаридами. Дисахариды состоят из 2-х одинаковых или различных моносахаридов. Различают дисахариды мальтозного типа связи, при этом молекула дисахарида образуется из двух молекул моносахаридов через кислородный мостик от гликозидного гидроксила одного моносахарида и гидроксила (обычно 4-го атома углерода) второго моносахарида. К таким моносахаридам относятся мальтоза, лактоза, целлобиоза. Для этих дисахаридов характерны все химические реакции, типичные для альдоз и кетоз, имеющих гликозидный гидроксил. Мальтоза – состоит из двух остатков α-D-глюкопиранозы, где атом углерода C-1 одной молекулы соединяется через кислородный мостик с 4-м атомом углерода второй молекулы. Мальтоза содержится в проросших зернах ячменя (солода), ржи, пшеницы, является промежуточным продуктом гидролиза крахмала, гликогена. Лактоза – молочный сахар, молекула образована из моносахаридов β-D-галактозы и α-D-глюкозы. Входит в состав гликопротеидов и гликолипидов, полисахаридов. 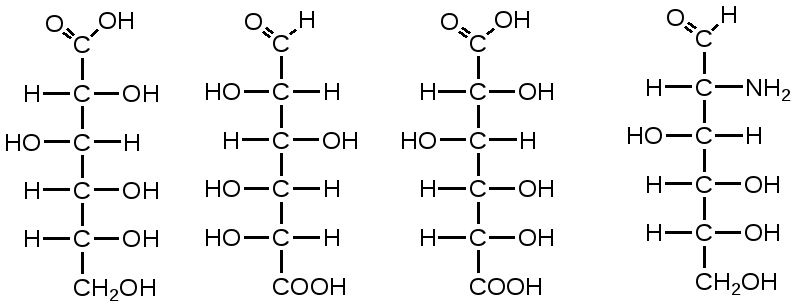 глюконовая глюкуроновая сахарная Д-глюкозамин кислота кислота кислота (хитозамин) 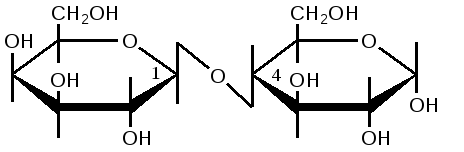 лактоза 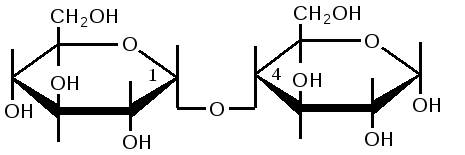 мальтоза 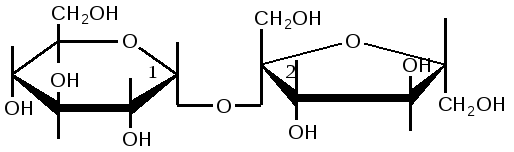 сахароза 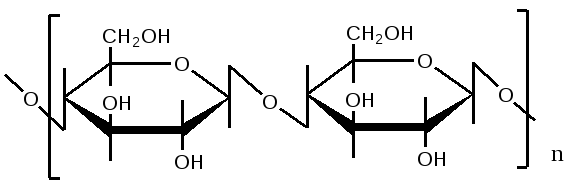 Участок молекулы целлюлозы Рис.8.2. Производные гексоз и дисахариды. Целлобиоза – дисахарид, образованный из остатков двух молекул (β-D-глюкозы, является промежуточным продуктом гидролиза клетчатки. Сахароза – свекловичный сахар, молекула состоит из остатков α-D-глюкозы и β-D-фруктозы, соединенных между собой кислородным мостиком, который образован за счет двух гликозидных гидроксилов (1,2), поэтому восстанавливающим свойством не обладает. При гидролизе молекула сахарозы расщепляется на глюкозу и фруктозу (рис.8.2.). Трегалоза – (грибной сахар) содержится в тканях грибов, водорослях, дрожжах, туберкулезной палочке, состоит из 2-х молекул глюкозы, соединенных гликозидной связью 1,1. Трисахариды. К ним относится раффиноза, состоящая из галактозы, глюкозы, фруктозы. Она содержится в свекловичном сахаре (мелассе), семенах хлопчатника. Полисахариды (гликаны) Различают: гомополисахариды – содержащие однотипные моносахариды и гетерополисахариды, содержащие разные моносахариды. Полисахариды, построенные из глюкозы называются глюканами (крахмал, гликоген), из маннозы – маннанами, из фруктозы – фруктозанами и т.д. Полисахариды являются основной массой растительного материала, важным продуктом в кормлении животных. Резервными полисахаридами являются – крахмал в тканях растений и гликоген – у животных. Г 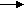 ликоген (полисахарид – из остатков D-глюкозы), соединенных между собой α (1 → 4)-гликозидными связями – откладывается в цитоплазме в виде гранул. Имеет разветвленную структуру (рис.8.3). Гликогены из различных тканей, а также различных видов животных имеют отличия по размеру молекул, содержат от 6 до 25 тысяч остатков глюкозы. Содержатся в печени и мышечной ткани. ликоген (полисахарид – из остатков D-глюкозы), соединенных между собой α (1 → 4)-гликозидными связями – откладывается в цитоплазме в виде гранул. Имеет разветвленную структуру (рис.8.3). Гликогены из различных тканей, а также различных видов животных имеют отличия по размеру молекул, содержат от 6 до 25 тысяч остатков глюкозы. Содержатся в печени и мышечной ткани.Крахмал – является запасным питательным веществом растений, построен из остатков D-глюкозы, соединенных между собой α (1 → 4) глюкозидными связями. Такая последовательность характерна для амилозы – компонента крахмального зерна, Молекулярная масса 16-97 кДа, количество остатков 100-600. 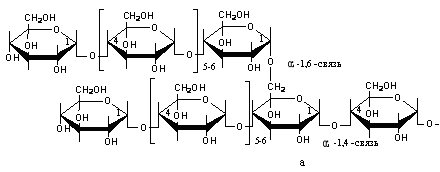 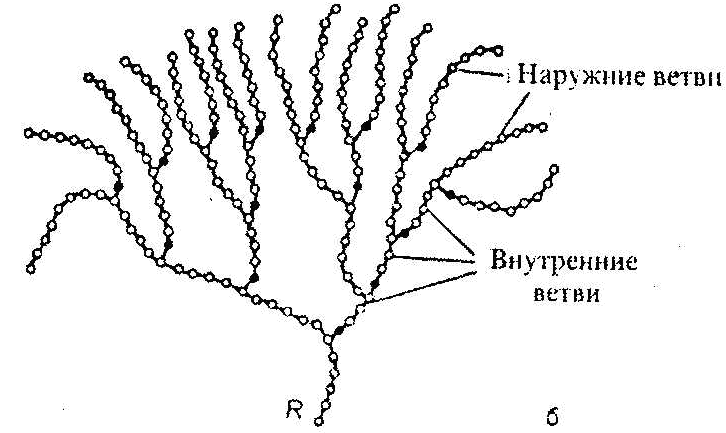 Рис.8.3. Строение отдельного участка (а) и всей (б) гликогена (по Майеру). Белые кружки – остатки глюкозы, соединенные α (1 → 4) связью; R -редуцирующая концевая группа. Внутренние цепи, или ветви, – участки между точками ветвления. Наружные цепи, или ветви, начинаются от точки ветвления и кончаются нередуцирующим остатком глюкозы. Амилопектин – другой компонент крахмального зерна, отличается характером ветвления, молекулярная масса колеблется в широких пределах от 48 кДа до 972 кДа, содержит 300-6000 глюкозных остатков. Ц еллюлоза (клетчатка) состоит из глюкозных остатков, соединенных β-(1 → 4)-глюкозидными связями. Целлюлоза – наиболее распространенное соединение углеводного характера. Молекулярная масса 972-1296 кДа, содержит 6000-8000 остатков β-глюкозы; в клеточной стенке находится в виде микрофибрилл.Ф руктозаны (леваны) – состоят из остатков D-фруктозы, соединенных β-(2 → 1) связями, являются резервными гомополисахаридами. Молекула инулина состоит примерно из 30 остатков молекул фруктозы.Декстраны состоят из остатков D-глюкозы, структурные единицы их остова связаны α-(1 → 6) связями. Декстраны – резервные полисахариды дрожжей и бактерий. Растворы декстранов имеют значительную вязкость. Маннаны – состоят из остатков маннозы, содержатся в растениях, дрожжах, бактериях; арабаны – из арабинозы, ксиланы – из ксилозы; это важные полисахариды, содержатся в кормовых растениях. Хитин – близок по строению к целлюлозе, состоит из остатков производного глюкозы. Широко распространенный полисахарид. Агар – состоит из D- и L- галактоз. Используется для приготовления питательных сред, в пищевой промышленности. Гетерополисахариды (гетерогликаны) Кроме целлюлозы в составе клеточных стенок находят гетерополисахарид гемицеллюлозу. В ее составе встречаются остатки галактозы, маннозы и т.д. Кроме того, встречаются гетерополисахариды, содержащие уроновые кислоты (слизи). Слизи выделяются при повреждении коры деревьев, содержатся в семенах льна. Основной компонент слизей – глюкуроновая и галактуроновая кислоты. Ряд гетерополисахаридов выделен из микроорганизмов, они определяют иммунологическую специфичность. Полиурониды (гликуронаны) – построены из уроновых кислот, к ним относятся пектины и альгиновые кислоты. Много пектина во фруктах. Пектины используются в пищевой и фармацевтической промышленности. Альгиновые кислоты - полимеры D-глюкуроной кислоты, содержатся в водорослях до 30% (сухого вещества растений). Мукополисахариды: широко распространена гиалуроновая кислота. Она содержится в соединительной ткани, в стекловидном теле глаза, синовиальной жидкости. Гиалуроновая кислота в синовиальной жидкости проявляет смазочные свойства. Гиалуроновая кислота состоит из D-глюкуроновой кислоты и 2-ацетамидо-2-дезокси-D-глюкозы (см. стр.79). Хондроитин – полисахарид, содержит D-галактозамин (в отличие от гиалуроновой кислоты). Входит в состав хондроитинсульфатов А, В, С – компонентов соединительной ткани, мукополисахаридов. Гепарин – антикоагулянт, является мукополисахаридом, молекулярная масса 10-20 кДа. Групповые вещества крови – относятся к гетерополисахаридам. На поверхности эритроцитов содержатся гетерополисахариды, связанные с белком. Этот гликопротеид мембраны эритроцитов состоит на 60% из углеводов, 40% из пептидов. Они определяют антигенные свойства крови. Обмен углеводов В организме животных имеется несколько десятков разных моносахаридов и много различных олиго- и полисахаридов. Функции углеводов заключаются в том, что они служат источником энергии, за счет их окисления обеспечивается около половины всей потребности животного в энергии, при этом главная роль принадлежит глюкозе и гликогену. Углеводы входят в состав структурно-функциональных компонентов клеток (пентозы нуклеотидов и нуклеиновых кислот, углеводы гликопротеинов и гликолипидов, гетерополисахариды межклеточного вещества). Из углеводов могут синтезироваться соединения других классов, в частности липиды и некоторые аминокислоты. Таким образом, углеводы выполняют разнообразные функции, важные для организма, но главная среди них – это энергетическая роль углеводов. В организме животных наиболее распространенным углеводом является глюкоза, которая выполняет роль связывающего звена между энергетическими и пластическими функциями углеводов; так, из нее могут образовываться все другие моносахариды и, наоборот, разные моносахариды могут превращаться в глюкозу. Общая схема обмена глюкозы представлена на рис. 8.4. Переваривание углеводов происходит под действием ферментов амилазы, целлюлазы, мальтазы, сахаразы, лактазы и целлобиазы.  Рис. 8.4. Общая схема метаболизма глюкозы: 1 - запасание углеводов в виде гликогена; 2 - мобилизация гликогена; 3 - 6 - анаболические превращения глюкозы; 7 - катаболизм глюкозы. Фермент амилаза гидролизует α-l-4-гликозидные связи крахмала с невосстанавливающегося конца молекулы (γ-амилаза в тканях животных) или без определенного порядка, образуя вначале олигосахариды, а затем мальтозы (α-амилаза в пищеварительном тракте). В ротовой полости распад крахмала начинается под действием α-амилазы и мальтазы слюны, в результате крахмал может распадаться до мальтозы и глюкозы, однако этих ферментов в слюне у животных содержится немного, пищевая масса находится в ротовой полости непродолжительное время. Дальнейший распад крахмала происходит в тонком отделе кишечника под действием α-амилазы поджелудочной железы. 1-6- гликозидные связи крахмала гидролизуются декстриназой. Мальтоза расщепляется мальтазой до 2-х молекул глюкозы, сахароза – сахаразой до глюкозы и фруктозы, лактоза – лактазой до глюкозы и галактозы. Считают, что мальтаза, лактаза, сахараза синтезируются в клетках стенки кишечника и гидролиз дисахаридов происходит на поверхности клеток или же внутри клеток кишечника. Клетчатка (целлюлоза) у жвачных животных гидролизуется под действием целлюлозолитических ферментов бактерий преджелудков, а у лошадей, кроликов – в толстом отделе кишечника (в слепой кишке). Целлюлаза бактериальных клеток гидролизует (3-гликозидные связи с образованием целлобиозы, а целлобиоза расщепляется под действием целлобиазы на 2 молекулы β-D-глюкозы, которая далее подвергается брожению под действием бактериальных ферментов с образованием летучих жирных кислот (ЛЖК) – пировиноградной, уксусной, молочной, масляной кислот и газов. ЛЖК всасываются через стенку рубца в кровь и служат энергетическим источником у жвачных – до 40 % энергетических потребностей организма удовлетворяются за их счет. Моносахариды всасываются из кишечника в кровь через клеточные мембраны путем облегченной диффузии с участием специальных переносчиков. Кроме того, существует активный транспорт с участием Na-K-AТФ-азы, которая обеспечивает перенос моносахаридов против градиента концентрации. Моносахариды после всасывания в кровь по воротной вене доставляются в печень. В клетках печени глюкоза фосфоролируется и ее избыток идет на синтез гликогена, который откладывается в печени и в мышечной ткани. Часть глюкозы поступает в большой круг кровообращения и доставляется во все ткани организма. Уровень сахара в крови поддерживается под влиянием гормонов поджелудочной железы инсулина (снижает уровень глюкозы) и глюкагона (мобилизует глюкозу из гликогена) в определенных пределах: у свиней 80-100 мг %, у лошадей 80-120 мг %, у крупного рогатого скота 40-60 мг %. Катаболизм глюкозы Расщепление глюкозы в тканях происходит аэробно – с участием кислорода – это основной путь катаболизма. Окисление глюкозы без доступа кислорода – анаэробное превращение, которое начинается с гликогена и заканчивается с образованием молочной кислоты называется гликогенолизом. Если этот процесс начинается с глюкозы, то называется гликолизом. Гликогенолиз Глюкоза, депонированная в форме гликогена, освобождается из него при участии фермента гликогенфосфорилазы. Этот фермент катализирует фосфорилирование 1,4-гликозидной связи невосстанавливающих концов гликогена. Глюкозо-1-фосфат, образующийся из гликогена, при участии фосфоглюкомутазы превращается в глюкозо-6-фосфат, то есть остаток фосфорной кислоты переносится на шестой углеродный атом глюкозы. Дальнейшие превращения глюкозо-6-фосфата такие же, как при гликолизе. Гликолиз – распад глюкозы начинается с фосфорилирования с участием АТФ [остаток фосфорной кислоты обозначается -PO-3H2 или (P)]. 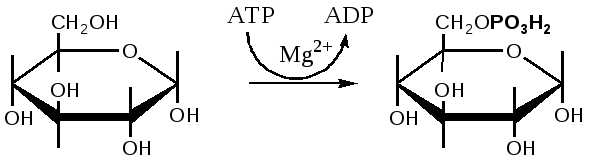 Глюкоза Глюкозо-6-фосфат В результате образуется глюкозо-6-фосфат. Фермент, осуществляющий эту реакцию – фосфогексокиназа (реже фосфоглюкокиназа). Следует подчеркнуть, что глюкоза способна проходить через клеточные мембраны, а глюкозо-6-фосфат не может, в результате фосфорилирования глюкозы она "запирается" в клетке. Глюкозо-6-фосфат, образующийся при гликолизе и гликогенолизе может подвергаться следующим обменным превращениям: а) образуется фруктозо-6-фосфат и происходят дальнейшие этапы гликолиза; б) происходит дефосфорилирование с образованием глюкозы и неорганического фосфата; в) вовлекается в пентозный путь распада. 1. Образование фруктозо-6-фосфата происходит в результате изомерного превращения с участием фермента фосфоглюкоизомеразы: 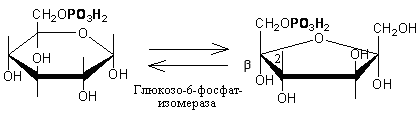 2. В следующей реакции происходит фосфорилирование фруктозо-6-фосфата с участием АТФ и фермента фосфофруктокиназы, в результате образуется фруктозо-1,6-ди-фосфат. 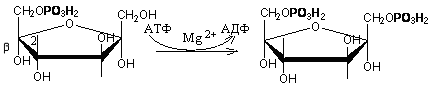 Фруктозо - 6 - фосфат Фруктозо -.1,6- дифосфат 3. Наличие двух остатков фосфата на противоположных концах молекулы гексозы приводит к резкому ослаблению связей между третьим и четвертым атомами углерода. В результате этого углевод легко расщепляется на 2 фосфотриозы под влиянием фермента альдолазы: 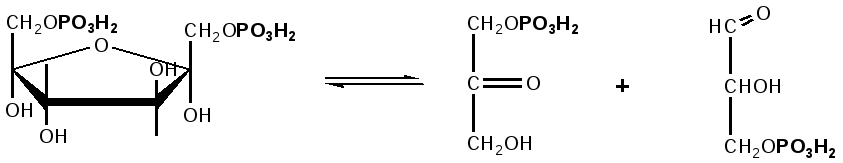 Фруктозо-1,6-дифосфат Диоксиацетон- Глицеральдегид фосфат - 3- фосфат При этом образуется 3-фосфоглицериновый альдегид (3%) и диоксиацетонфосфат (97%), который превращается в 3-фосфоглицериновый альдегид: 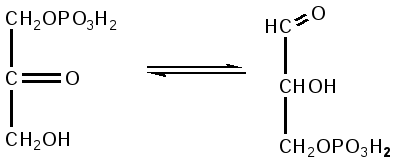 Диоксиацетонфосфат Глицеральдегид-3-фосфат На этом первый этап гликолиза завершается. На втором этапе происходит окисление 3-фосфоглицеринового альдегида до пировиноградной кислоты. 4. 3-фосфоглицериновый альдегид под влиянием фермента глицеральдегиддигидрогеназы и НАД вступает в реакцию образования 1,3-дифосфоглицериновой кислоты: 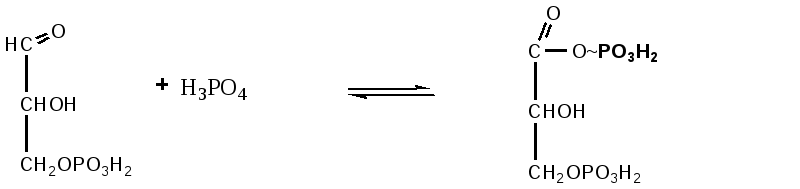 D-глицеральдегид- 1,3-дифосфоглицериновая 3-фосфат кислота При этом в начале реакции образуется фермент-субстратный комплекс, который в присутствии фосфорной кислоты распадается с образованием нативного HS-фермента (глицеральдегид-3-фосфатдегидрогеназы) и 1,3-дифосфоглицериновой кислоты. При этом энергия окисления 3-фосфоглицеринового альдегида не рассеивается в виде тепла, а используется для образования богатых энергией фосфорных связей 1,3-дифосфоглицериновой кислоты. Богатые энергией (макроэргические) связи в формуле обозначаются в виде « |
Реакция переноса остатка фосфорной кислоты с богатой энергией связью 1,3-дифосфоглицериновой кислоты на АДФ, в результате образуется АТФ. Реакция происходит с участием фермента фосфоглицераткиназы:
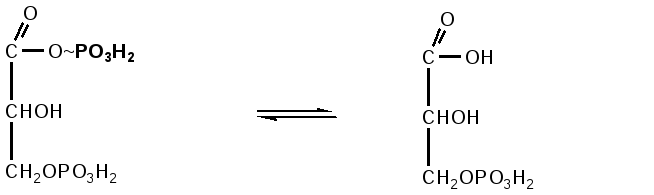

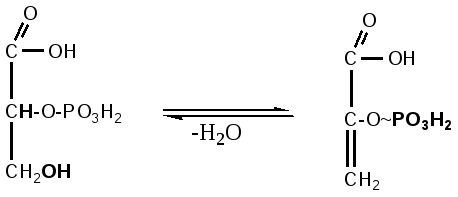
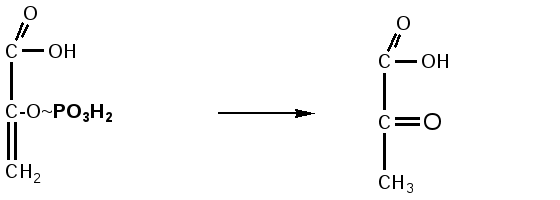
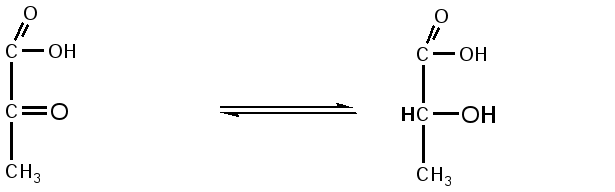
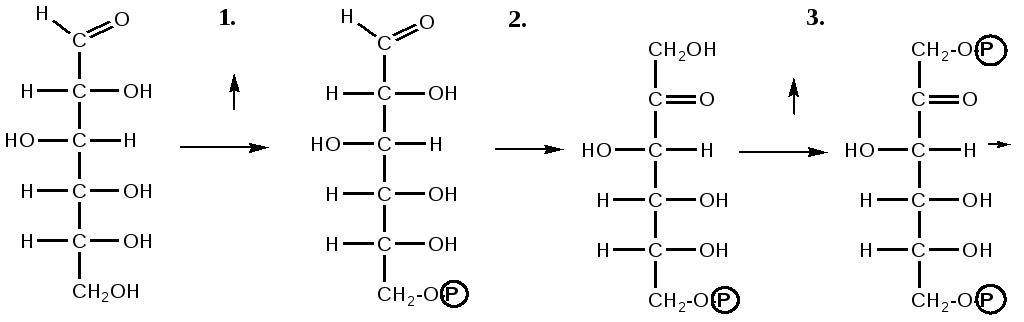 глюкоза глюкозо- фруктозо-6-фосфат фруктозо-
глюкоза глюкозо- фруктозо-6-фосфат фруктозо-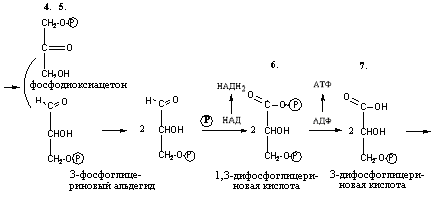
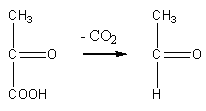
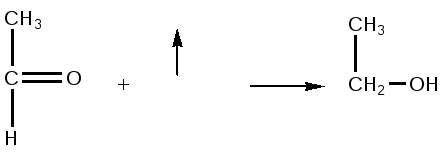
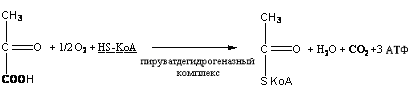
Реакция окислительного декарбоксилирования подавляется молекулами АТФ, то есть накопление молекул АТФ является сигналом для прекращения реакции декарбоксилирования пировиноградной кислоты.
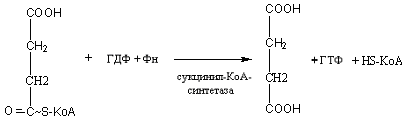
Цикл трикарбоновых кислот или цикл Кребса является завершающим этапом распада энергетических материалов в клетке. В нем происходит окисление активированной уксусной кислоты (ацетил
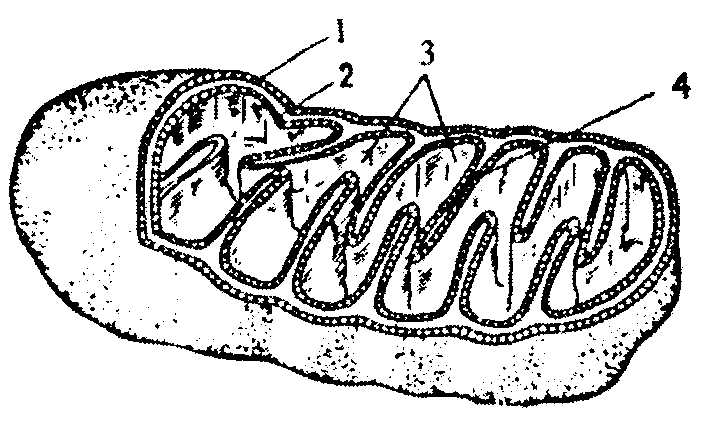

 ТФ + АДФ АТФ + ГДФ
ТФ + АДФ АТФ + ГДФ