биохимия хазипов. Биохимия животных Электронный дидактический комплекс (эдк)
 Скачать 14.38 Mb. Скачать 14.38 Mb.
|

11. БИОСИНТЕЗ БЕЛКАБиосинтез белка является сложным процессом, происходящим во всех клетках, органах и тканях организма. Наибольшее количество белка синтезируется в печени. Белоксинтезирующая система включает около 300 соединений, в том числе:
Рибосомы – рибонуклеопротеиды, состоящие на 50-65 % из рибосомальной РНК и белка – 35-50 %. Известно, что рибосомы животных клеток (эукариотов) в 2 раза крупнее (80 S) рибосом бактерий (70 S) (прокариот). Рибосомы состоят из двух частиц, 60 S и 40 S (рибосомы животных клеток – 80 S) и 50 S и 30 S (рибосомы бактерий – 70 S). Количество рибосом в одной клетке составляет приблизительно 100000. В присутствии ионов Mg2+ рибосомы распадаются на две части (60 и 40 S). Рибосома животных клеток содержит 3 вида РНК – 28, 18 и 5S и свыше 70 различных белков. Общую схему строения рибосом животных клеток можно представить в следующем виде:  РНК рибосом синтезируется на матрице ДНК в ядре. Рибосомные белки имеют цитоплазматическое происхождение, затем они транспортируются в ядрышки, где происходит формирование рибосомных субчастиц. После этого субчастицы переходят в цитоплазму и несколько рибосом (до 50-80 рибосом) вместе с информационной РНК образуют полисомы или полирибосомы - место синтеза белка: Аминоацил-тРНК-синтетазы – это ферменты с молекулярной массой (≈) 100000 Да, обеспечивающие активирование аминокислот и связывание их с т-РНК. Эти ферменты абсолютно специфичны, то есть узнают только одну какую-либо аминокислоту (а) и т-РНК. Считают, что этот фермент имеет три активных центра связывания: для аминокислоты, т-РНК и АТФ. Количество этих ферментов должно быть не менее 20 – для каждой аминокислоты отдельно. Транспортные РНК. На долю т-РНК приходится 10-15 % из общего количества РНК, для каждой аминокислоты имеется своя т-РНК, а для некоторых аминокислот по несколько т-РНК, например для серина – 5, лизина и глицина по 4 разных т-РНК. Молекулярная масса большинства т-РНК колеблется в пределах 24000-29000 Да, они содержат от 75 до 85 нуклеотидов. т-РНК имеют сходную структуру, с тремя участками связывания: на 3'-гидроксильном конце у всех т-РНК имеется триплет ЦЦА- ОН, к которому через 3'-OH присоединяется специфическая аминокислота. Следующий связывающий участок – антикодон из трех нуклеотидов. Антикодон комплементарен кодону и-РНК. Например,  5'- концевой нуклеотид во всех т-РНК это ГМФ со свободной 5'-фосфатной группой (рис. 11.1.).  основание Рис.11.1. Обобщенная схема вторичной структуры т-РНК, представленной в форме клеверного листа. Черные кружки соответствуют основаниям в спаренных областях, связанным водородными связями; светлые кружки соответствуют неспаренным основаниям. R - пуриновый нуклеозид; Y - пиримидиновый нуклеозид. Функция т-РНК – специфическое связывание каждой т-РНК со своей аминокислотой. и-РНК (информационная РНК) – переносит информацию для синтеза белка от ДНК к рибосоме, служит матрицей, на которой происходит синтез белка. и-РНК синтезируется на ДНК по правилу комплементарности, является копией участка ДНК (гена, цистрона). Размер молекулы зависит от размера белка. Молекула и-РНК образуется в виде предшественника, затем происходит созревание, то есть удаление участков интронов, не входящих в состав зрелой молекулы:  Участки экзонов сшиваются в процессе созревания и-РНК. Полиадениловый хвост на 3'-конце обеспечивает стабильность иРНК. и-РНК комплементарна определенному участку молекулы ДНК, например: 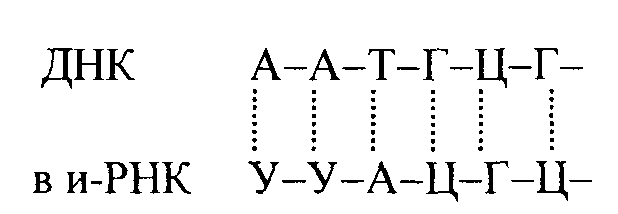 Генетический код. Генетическая информация закодирована в ДНК в виде последовательности нуклеотидов и эта последовательность переводится в нуклеотидную потельность и-РНК. Код является триплетным, то есть информация для каждой аминокислоты состоит из трех нуклеотидов. Например, для включения в белковую молекулу фенилаланина код содержит УУУ- триплет из молекул уридиловой кислоты (таблица 11.1.). Генетический код является вырожденным, то есть для кодирования одной аминокислоты имеется несколько кодонов. Код является непрерывным, то есть отсутствуют знаки, сигналы, обозначающие начало и конец кода. Например: АЦЦЦТГААЦ и т.д. Код является универсальным – для всех живых систем от E.Coli до человека. Из мыслимых 64 кодонов 61 имеет смысл, то есть кодируют определенную аминокислоту, а кодоны УАГ, УАА, УГА кодируют терминацию. Таблица 11.1. Генетический код Второй нуклеотид кодона  Этапы синтеза белка В процесс синтеза белка включается около 300 типов макромолекул – белков и нуклеиновых кислот, из них около 100 участвуют в активировании аминокислот и переносе их на рибосомы, свыше 60 – в составе рибосом, свыше 10 белковых факторов участвуют в процессах трансляции. Синтез идет в направлении от свободного аминного конца к свободному карбоксильному концу: NH2 → СООН (Рис. 11.2).  Рис. 11.2. Принципиальная схема биосинтеза белка (по А.С. Спирину). Активирование аминокислот происходит в цитоплазме при помощи ферментов аминоацил-т-РНК-синтетаз с участием АТФ, в результате образуется аминоацил-т-РНК:  Активированная аминокислота переносится к месту синтеза белка – к полирибосомному комплексу. Следующий этап биосинтеза белка – трансляция – происходит в рибосоме и имеет 3 стадии: инициацию, элонгацию, терминацию. Инициация трансляции обеспечивается специальными белками, их около 10, начальный участок содержит метионин, то есть начало синтеза обусловлено кодоном формил-метионина (начальный метионин). При этом антикодон (мет-т-РНК) ЦАЦ соединяется с кодоном АУГ и-РНК, образуется инициативный комплекс – к 40 S рибосомной части присоединяется мет-тРНК, затем 60 S субчастица рибосомы(рис.11.3.).  Рис.11.3. Схематическое изображение взаимодействия формилметионил-т-РНК и иРНК с 40S субчастицей рибосомы (а) и транслирующей 80 S рибосомы (б). После образования комплекса 80 S рибосома + и-РНК, начинается элонгация, то есть удлинение полипептидной цепи. В рибосоме образуется пептидная связь между аминокислотами за счет энергии ГТФ. После присоединения очередной аминокислоты происходит движение рибосомы вдоль цепи и-РНК с 5' к 3' концу на 3 нуклеотида (один ко дон) для каждой аминокислоты. Терминация трансляции осуществляется с участием трех белковых факторов. Последний терминирующий кодон обуславливает отделение и-РНК от 80S и 80S рибосома распадается на составные субчастицы 60 и 40 S, которые поступают в клеточный пул и используются для образования и-РНК – рибосомного комплекса. Одна и-РНК транслируется не одной рибосомой, а одновременно многими рибосомами (полирибосомы, полисомы), что значительно ускоряет синтез белка (рис. 11.2.). Постсинтетическая модификация белков происходит в несколько этапов: в начале удаляется сигнальный пептид (из 15-30 аминокислотных остатков) – специфическая протеаза отщепляет начальный N - концевой метионин. Таким образом, отщепляется часть молекулы у проферментов (пепсиноген, проинсулин и т.д.). Затем происходит химическая модификация ферментных белков -ковалентное присоединение простетической группы к молекуле белка, например, присоединение пиридоксальсульфата к белковой части аминотрансфераз; гликозилирование – присоединение олигосахаридных остатков в гликопротеинах; гидроксилирование пролина, лизина, метилирование лизина и глутамата, ацетилирование N-концевых аминокислот, фосфорилирование серина, треонина, тирозина и т.д. Укладка белковой молекулы -образование вторичной, третичной и четвертичной структур так же происходит после синтеза – образования первичной структуры белковой молекулы. Мультиферментный механизм синтеза белка Описанный выше матричный механизм синтеза белков лежит в основе синтеза почти всех белков живых организмов. Но синтез ряда низкомолекулярных пептидов может происходить без нуклеиновых кислот и рибосом. В настоящее время доказано, что синтез природных антибиотиков-пептидов: грамицидина S и тироцидина происходит таким способом. Белки, экстрагированные из бактериальных клеток Вас. brevis обеспечивают точность сборки циклического пептида грамицидина S, состоящего из 10 аминокислотных остатков. Антибиотик тироцидин и 13 членный циклический пептид микобациллин синтезируется подобным образом. |