Биология экзамен. Биология наука о жизни, об общих закономерностях существования и развития живых существ. Место и задачи предмета в системе медицинского образования. Биология
 Скачать 0.56 Mb. Скачать 0.56 Mb.
|

верхности(от англ, external) - наружной, более близкой к внешней среде поверхности скола (внутренней поверхности наружного листка).Билеты по разделам! 1.Биология – наука о жизни, об общих закономерностях существования и развития живых существ. Место и задачи предмета в системе медицинского образования. Биология — наука о жизни (от греч. биос — жизнь, логос — наука) — изучает закономерности жизни и развития живых существ. Термин «биология» был предложен немецким ботаником Г.Р. Тревиранусом и французским естество- испытателем Ж.-Б. Ламарком в 1802 году независимо друг от друга. Биология относится к естественным наукам. Разделы науки биологии можно классифицировать по-разному. Например, в биологии выделяют науки по объектам исследования: о животных — зоологию; о растениях — ботанику; анатомию и физиологию человека как основу медицинской науки. В пределах каждой из этих наук имеются более узкие дисциплины. Например, в зоологии выделяют протозоологию, энтомологию, гельминтологию и другие. Биологию классифицируют по дисциплинам, изучающим морфологию (строение) и физиологию (функции) организмов. К морфологическим наукам относят, например, цитологию, гистологию, анатомию. Физиологические науки — это физиология растений, животных и человека. Для современной биологии характерно комплексное взаимодействие с другими науками (химией, физикой, математикой) и появление новых сложных дисциплин. Значение биологии для медицины велико. Биология — теоретическая основа медицины. Врач древней Греции Гиппократ (460-274 г. до н.э.) считал, что «необходимо, чтобы каждый врач понимал природу». Во всех теоретических и практических медицинских науках используются общебиологические обобщения. Теоретические исследования, проводимые в различных областях биологии, позволяют использовать полученные данные в практической деятельности медицинских работников. Например, открытие структуры вирусов, возбудителей инфекционных заболеваний (оспы, кори, гриппа и других), и способов их передачи, позволило ученым создать вакцину, предотвращающую распространение этих заболеваний или снижающую риск гибели людей от этих тяжелых инфекций. 2.Сущность жизни. Основные свойства и уровни организации живой материи. Согласно определению, данному ученым-биологом М.В. Волькенштейном (1965 г.), «живые организмы представляют собой открытые, саморегулирующиеся, самовоспроизводящиеся системы, построенные из биополимеров — белков и нуклеиновых кислот». Через живые открытые системы проходят потоки энергии, информации, вещества. Живые организмы отличаются от неживых признаками, совокупность которых определяет их жизненные проявления. СВОЙСТВА ЖИВОГО 1. Химический состав. Живые существа состоят из тех же химических элементов, что и неживые, но в организмах есть молекулы веществ, характерных только для живого (нуклеиновые кислоты, белки, липиды). 2. Дискретность и целостность. Любая биологическая система (клетка, организм, вид и т.д.) состоит из отдельных частей, т.е. дискретна. Взаимодействие этих частей образует целостную систему (например, в состав организма входят отдельные органы, связанные структурно и функционально в единое целое). 3. Структурная организация. Живые системы способны создавать порядок из хаотичного движения молекул, образуя определенные структуры. Для живого характерна упорядоченность в пространстве и времени. Это комплекс сложных саморегулирующихся процессов обмена веществ, протекающих в строго определенном порядке, направленном на поддержание постоянства внутренней среды — гомеостаза. 4. Обмен веществ и энергии. Живые организмы — открытые системы, совершающие постоянный обмен веществом и энергией с окружающей средой. При изменении условий среды происходит саморегуляция жизненных процессов по принципу обратной связи, направленная на восстановление постоянства внутренней среды — гомеостаза. Например, продукты жизнедеятельности могут оказывать сильное и строго специфическое тормозящее воздействие на те ферменты, которые составили начальное звено в длинной цепи реакций. 5. Самовоспроизведение. Самообновление. Время существования любой биологической системы ограничено. Для поддержания жизни происходит процесс самовоспроизведения, связанный с образованием новых молекул и структур, несущих генетическую информацию, находящуюся в молекулах ДНК. 6. Наследственность. Молекула ДНК способна хранить, передавать наследственную информацию, благодаря матричному принципу репликации, обеспечивая материальную преемственность между поколениями. 7. Изменчивость. При передаче наследственной информации иногда возникают различные отклонения, приводящие к изменению признаков и свойств у потомков. Если эти изменения благоприятствуют жизни, они могут закрепиться отбором. 8. Рост и развитие. Организмы наследуют определенную генетическую информацию о возможности развития тех или иных признаков. Реализация информации происходит во время индивидуального развития — онтогенеза. Наопределенном этапе онтогенеза осуществляется рост организма, связанный с репродукцией молекул, клеток и других биологических структур. Рост сопровождается развитием. 9. Раздражимость и движение. Все живое избирательно реагирует на внешние воздействия специфическими реакциями благодаря свойству раздражимости. Организмы отвечают на воздействие движением. Проявление формы движения зависит от структуры организма. Уровни организации живой материи1. Молекулярный – это уровень сложных органических веществ – белков и нуклеиновых кислот. На этом уровне происходят химические реакции обмена веществ (гликолиз, кроссинговер и т.п.), но молекулы сами по себе еще не могут считаться живыми. 2. Клеточный. На этом уровне возникает жизнь, потому что клетка – минимальная единица, обладающая всеми свойствами живого. 3. Органно-тканевой – характерен только для многоклеточных организмов. 4. Организменный – за счет нервно-гуморальной регуляции и обмена веществ на этом уровне осуществляется гомеостаз, т.е. сохранение постоянства внутренней среды организма. 5. Популяционно-видовой. На этом уровне происходит эволюция, т.е. изменение организмов, связанное с приспособлением их к среде обитания под действием естественного отбора. Наименьшей единицей эволюции является популяция. 6. Биогеоценотический (совокупность популяций разных видов, связанных между собой и окружающей неживой природой). На этом уровне происходит круговорот веществ и превращение энергии, а также саморегуляция, за счет которой поддерживается устойчивость экосистем и биогеоценозов 7. Биосферный. На этом уровне происходит глобальный круговорот веществ и превращение энергии, а так же взаимодействие живого и неживого вещества планеты. Раздел 1 Молекулярно-генетический уровень. 1.Химическая организация генетического материала. Строение нуклеиновых кислот. Генетический код. Материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя. Нуклеиновые кислоты являются макромолекулами, т.е. отличаются большой молекулярной массой. Это полимеры, состоящие из мономеров — нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1' присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5' с помощью эфирной связи — фосфат; у третьего атома углерода С-3' всегда имеется гидроксильная группа — ОН (рис. 3.1). Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь. В результате образуется полинуклеотидная цепь. Остов цепи состоит из чередующихся молекул фосфата и сахара. К молекулам пентозы в положении С-1' присоединено одно из перечисленных выше азотистых оснований. Среди нуклеиновых кислот различают два вида соединений: дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (РНК) кислоты. Изучение состава основных носителей наследственного материала — хромосом — обнаружило, что их наиболее химически устойчивым компонентом является ДНК, которая представляет собой субстрат наследственности и изменчивости. ДНК состоит из нуклеотидов, в состав которых входят сахар — дезоксирибоза, фосфат и одно из азотистых оснований — пурин (аденин или гуанин) либо пиримидин (тимин или цитозин). Особенностью структурной организации ДНК является то, что ее молекулы включают две полинуклеотидные цепи, связанные между собой определенным образом. В соответствии с трехмерной моделью ДНК, предложенной в 1953 г. американским биофизиком Дж. Уотсоном и английским биофизиком и генетиком Ф. Криком, эти цепи соединяются друг с другом водородными связями между их азотистыми основаниями по принципу комплементарности. Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи, а между гуанином и цитозином разных цепей образуются три водородные связи. Такое соединение азотистых оснований обеспечивает прочную связь двух цепей и сохранение равного расстояния между ними на всем протяжении. Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5'-конец одной цепи соединяется с 3'-концом другой, и наоборот. Таким образом, в структурной организации молекулы ДНК можно выделить первичную структуру —полинуклеотидную цепь, вторичную структуру—две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру — трехмерную спираль с приведенными выше пространственными характеристиками. Роль РНК в реализации наследственной информации В отличие от молекул ДНК рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований — аденин, гуанин, урацил или цитозин. РНК синтезируется на молекулах ДНК при помощи ферментов РНК-полимераз с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Все многообразие РНК, действующих в клетке, можно разделить на три основных вида: мРНК, тРНК, рРНК. Матричная, или информационная, РНК (мРНК, или иРНК). Транскрипция. Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает «инструкция» о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией. Транспортная РНК (тРНК). Трансляция. Доставляя необходимые аминокислоты к месту сборки пептидных цепей, тРНК выполняет функцию трансляционного посредника. Наряду с функцией точного узнавания определенного кодона в мРНК молекула тРНК осуществляет доставку к месту синтеза пептидной цепи строго определенной аминокислоты, зашифрованной с помощью данного кодона. Специфическое соединение тРНК со «своей» аминокислотой протекает в два этапа и приводит к образованию соединения, называемого аминоацил-тРНК. Рибосомная РНК (рРНК). Рибосомный цикл синтеза белка. Процесс взаимодействия мРНК и тРНК, обеспечивающий трансляцию информации с языка нуклеотидов на язык аминокислот, осуществляется на рибосомах. Последние представляют собой сложные комплексы рРНК и разнообразных белков, в которых первые образуют каркас. Рибосомные РНК являются не только структурным компонентом рибосом, но и обеспечивают связывание их с определенной нуклеотидной последовательностью мРНК. В ходе трансляции можно выделить три фазы: инициацию, элонгацию и терминацию синтеза пептидной цепи. Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК (рис. 3.32). Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном. Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы. 2.Ген – функциональная единица наследственности. Особенности структурной организации гена. Функциональные группы генов. Свойства генов. Ген - функционально активная единица ДНК, предназначенная для синтеза той или иной молекулы РНК, в т.ч. кодирующая пептид, либо выполняющая регуляторную функцию. Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар нуклеотидов, в которых закодирована генетическая информация о первичной структуре белка (число и последовательность аминокислот). Три подряд расположенных нуклеотида представляют собой кодон, который и определяет, какая аминокислота будет располагаться в данной позиции в белке. Доказательствами хромосомной локализации генов явились: открытие генов, наследующихся сцеплено с полом (локализация генов в половых хромосомах, X или Y); сцепленное наследование группы признаков. Было показано наличие определенного числа групп сцепления генов, соответственно гаплоидному числу хромосом конкретного биологического вида. Кроме того, были получены генетические и цитологические доказательства кроссинговера - обмена генами между гомологичными хромосомами, приводящего к рекомбинации генов. Величина генетической рекомбинации (процент кроссинговера-перекреста) отражает расстояние между генами одной группы сцепления: чем дальше отстоят друг от друга гены, тем больше процент кроссинговера. Таким образом, было доказано, что гены в хромосоме располагаются в линейном порядке, и каждый ген имеет свое определенное местоположение - локус. Классификация генов 1) По месту локализации генов в структурах клетки различают расположенные в хромосомах ядра ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями. 2) По месту локализации гена в хромосомах: аллельные и неаллельные 3) По функциональному значению различают структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены -- последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.). 4) По влиянию на физиологические процессы в клетке различают летальные, условно летальные, гены-мутаторы и гены-антимутаторы, онкогены и антионкогены. 5) Гены “домашнего хозяйства” и роскоши 3. Понятие об опероне. Регуляция активности структурных генов. Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов. Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб и Моно, за что получили Нобелевскую премию в 1965 году. Опероны по количеству цистронов делят на моно-, олиго- и полицистронные. Характерным примером оперонной организации генома прокариот является лактозный оперон. Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор. Регуляция экспрессии генов у прокариот Изучение регуляции генной активности у прокариот привело французских микробиологов Ф. Жакоба и Ж. Моно к созданию (1961) оперонной модели регуляции транскрипции. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. Например, это могут быть гены, которые детерминируют синтез ферментов, участвующих в метаболизме какого-либо вещества или в синтезе какого-то компонента клетки. Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор. Особенностью прокариот является транскрибирование мРНК со всех струтурных генов оперона в виде одного полицистронного транскрипта, с которого в дальнейшем синтезируются отдельные пептиды. Примером участия генетических и негенетических факторов в регуляции экспрессии генов у прокариот может служить функционирование лактозного оперона у кишечной палочки Е. colt . Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, РНК-полимераза взаимодействует с промотором и осуществляет транскрипцию полицистронной мРНК. Последняя обеспечивает синтез сразу всех ферментов, участвующих в метаболизме лактозы. Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к восстановлению способности репрессора соединяться с оператором и прекращению транскрипции генов Z, Y, А. Таким образом, регуляция экспрессии генов, организованных у прокариот в опероны, является координированной. Синтез полицистронной мРНК обеспечивает одинаковый уровень синтеза всех ферментов, участвующих в биохимическом процессе. Регуляция экспрессии генов у эукариот У эукариот не установлено оперонной организации генов. Гены, определяющие синтез ферментов одной цепи биохимических реакций, могут быть рассеяны в геноме и, очевидно, не имеют, как у прокариот, единой регулирующей системы (ген-регулятор, оператор, промотор). В связи с этим синтезируемые мРНК у эукариот моноцистронны, т.е. являются матрицами для отдельных пептидных цепей. У многих эукариотических генов, кодирующих белки и транскрибируемых РНК-полимеразой II, в ДНК имеется несколько областей, которые узнаются разными белками-регуляторами. Одной из них является область, расположенная вблизи промотора. Она включает около 100 пар нуклеотидов, в том числе ТАТА-блок, располагающийся на расстоянии 25 пар нуклеотидов от точки начала транскрипции. Установлено, что для успешного присоединения РНК-полимеразы II к промотору необходимо предварительное соединение с ТАТА-блоком особого белка — фактора транскрипции — с образованием стабильного транскрипционного комплекса. Именно этот комплекс ДНК с белком узнается РНК-полимеразой II. Последовательности нуклеотидов, примыкающие к ТАТА-блоку, формируют требуемый для транскрипции элемент, расположенный перед промотором. Другая область, играющая важную роль в регуляции активности эукариотических генов, располагается на большом расстоянии от промотора (до нескольких тысяч пар нуклеотидов) и называется энхансером (от англ. enhance —усиливать). Особенностью регуляции экспрессии эукариотических генов является также существование белков-регуляторов, которые способны контролировать транскрипцию многих генов, кодирующих, возможно, другие белки-регуляторы. В связи с этим некоторые (главные) белки-регуляторы обладают координирующим влиянием на активность многих генов и их действие характеризуется плейотропным эффектом (рис. 3.88). Примером может служить существование белка, который активирует транскрипцию нескольких специфических генов, определяющих дифференцировку предшественников жировых клеток. Следующая особенность регуляции генной активности у эукариот связана с образованием стойкого комплекса ДНК с белками — хроматина (см. разд. 3.5.2.2). Ведущая роль в компактизации ДНК принадлежит гистонам, поэтому они, несомненно, участвуют и в процессах регуляции генной активности (см. разд. 3.5.4). Непременным условием для осуществления транскрипции у эукариот является предварительная декомпактизация хроматина на соответствующем участке, где временно утрачивается связь с Hi-гистонами и несколько ослабляется связь с нуклеосомными гистонами. Правда, нуклеосомная организация хроматина не утрачивается даже в ходе транскрипции, однако контакт ДНК и негистоновых белков становится возможным и происходит дерепрессия гена. Наконец, регуляция процесса реализации наследственной информации может осуществляться и на стадии посттрансляционных изменений. Прекращение этих процессов обусловливает задержку в формировании активных молекул белка при наличии необходимых для этого пептидных цепей. Например, для формирования активной формы белкового гормона — инсулина — из проинсулина должны вырезаться две субъединицы. Торможение этих процессов уменьшает выход конечного активного продукта. Таким образом, рассмотренный выше пример регуляции экспрессии генов демонстрирует сложнейшие взаимосвязи, которые существуют между ними в геноме. Формирование любого признака поэтому нельзя рассматривать как результат действия одной пары аллельных генов в генотипе. В любом случае регуляция экспрессии ответственного за этот признак гена осуществляется при участии других генов. 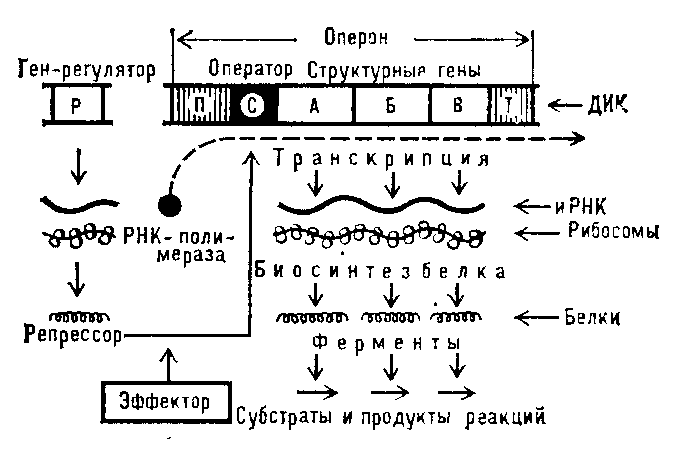 4. Кодирование и реализация наследственной информации. Этапы биосинтеза белка (транскрипция, посттранскрипционные процессы, трансляция, посттрансляционные процессы). СИНТЕЗ БЕЛКА Биосинтез белков идет в каждой живой клетке. Наследственная информация хранится в молекулах ДНК, а передается в цитоплазму информационной РНК (иРНК), которая комплементарна одной нити молекулы ДНК. Отрезок ДНК, содержащий информацию о структуре одного белка, называют геном. Транскрипцией называется перенос информации с ДНК на РНК. Матрицей для синтеза РНК служит только одна из двух цепей молекулы ДНК. Данный процесс происходит в ядре по принципу комплементарности. В транскрипции различают 4 стадии: 1. Связывание РНК-полимеразы (фермента, осуществляющего синтез РНК) с промотором. 2. Инициация – начало синтеза иРНК. В реакции участвуют нуклеотидтрифосфаты. 3. Элонгация – рост цепи РНК. Для этого процесса необходимы ионы магния. 4. Терминация – завершение синтеза РНК в участках-терминаторах. Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации. Трансляцией называется процесс синтеза белка на рибосомах, направляемый матрицей иРНК. Стадии: 1. Стадия активации аминокислот. Каждая аминокислота взаимодействует с молекулой АТФ под действием специфичного для каждой аминокислоты фермента - кодазы. 2. Присоединение фосфорилированных аминокислот к тРНК с образованием комплекса. Фермент при этом освобождается. 3. Собственно трансляция, или поликонденсация, аминокислотных остатков с образованием пептидных связей. 4. Терминация ПОСТТРАНСКРИПЦИОННЫЕ И ПОСТТРАНСЛЯЦИОННЫЕ ПРОЦЕССЫ Внутри самого структурного гена также различают участки смысловые (экзоны) и несмысловые, «молчащие» (интроны). На матрице структурного гена синтезируется так называемая про-иРНК – копия всего гена. Затем про-иРНК подвергается созреванию – процессингу – в ходе которого там же в ядре все несмысловые участки вырезаются, а концы кодирующих последовательностей соединяются. Процесс соединения называется сплайсингом. Все интроны вырезаются не всегда. При изменении условий часть из них может остаться в зрелой иРНК. Иногда же вырезаются какие-либо экзоны. Таким образом, один ген способен кодировать структуру нескольких белков. По окончании трансляции первая аминокислота в белковой цепи удаляется. Полученный полипептид имеет только первичную структуру, чтобы он приобрел функциональную активность, по выходу из рибосомы синтезируется вторичная, третичная, а у некоторых белков – четвертичная структура. РЕГУЛЯЦИЯ БИОСИНТЕЗА БЕЛКА Регуляцию процессов биосинтеза белков описали в 1961 г. описали французские ученые Франсуа Жакоб, Андре-Мишель Львов, Жан Люсьен Моно у бактерий. У прокариот можно выделить структуру, называемую опероном. Это участок ДНК, состоящий из следующих частей: Промотор – определенная последовательность нуклеотидов, которая связывается с ферментом РНК-полимеразой. Оператор – участок ДНК, связанный с белком-репрессором. До тех пор, пока эта связь не нарушена, РНК-полимераза не будет считывать информацию. Ряд структурных генов, кодирующих информацию обо всех ферментах, участвующих в расщеплении определенного субстрата или синтезе какого-либо вещества. Терминатор – участок ДНК, на котором заканчивается считывание. При поступлении субстрата в клетку его молекулы связываются с белком-репрессором, который после этого теряет способность взаимодействовать с оператором, происходит инициация, а РНК-полимераза начинает синтезировать иРНК, комплементарную структурным генам (элонгация). Затем в рибосомах синтезируются соответствующие ферменты, расщепляющие данный субстрат. Если субстрата не остается, освобождается белок-репрессор, который вновь блокирует оператор, и синтез иРНК и ферментов прекращается. Таким образом, бактерия синтезирует только те ферменты, которые ей необходимы в данный момент, что позволяет экономить энергию. У эукариот регуляция активности генов значительно сложнее. На нее влияют гормоны, медиаторы, другие биологически активные вещества, причем транскрипция, выход иРНК и трансляция регулируются отдельно и могут быть разделены во времени. Кроме того, структурные гены, кодирующие белки, необходимые для выполнения одной функции, могут быть расположены в разных хромосомах. Структура самих генов эукариот также более сложная. У эукариот собственно гены разделены участками «молчащей», нетранскрибируемой ДНК – спейсерами. 5.Репликация ДНК: принципы, ферментативные системы, механизмы контроля репликации ДНК. Значение репликации ДНК в ходе онтогенеза и при смене поколений. Репликация ДНК – это процесс, в результате которого из одной молекулы образуются две дочерние, полностью идентичные материнской, что обеспечивает передачу наследственной информации от поколения к поколению. Репликация осуществляется в соответствии со следующими принципами: Комплементарности; Полуконсервативности, т.е. каждая дочерняя спираль включает в себя одну синтезированную и одну материнскую полинуклеотидные цепи; Антипараллельности. ДНК-полимераза движется от 3′ к 5′-концу, т.о. дочерняя нить подстраивается антипараллельно, т.е. от 5′ к 3′-концу. Прерывистости. Синтез дочерних нитей ДНК начинается сразу в нескольких местах. Вся молекула не раскручивается, т.к. она очень велика. Участок между двумя точками, где начинается синтез дочерних цепей, называется репликоном. В каждом репликоне есть так называемая вилка репликации, т.е. та часть молекулы, которая под действием специальных ферментов уже расплелась и служит матрицей для синтеза. В процессе репликации принимает участие много белков-ферментов. ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи. ДНК-топоизомераза разрывает связь между остатками фосфорной кислоты и дезоксирибозы в одной из полинуклеотидных цепей ДНК, что дает ей возможность вращаться вокруг второй и снимать напряжение, вызываемое расплетением спирали и расхождением цепей в репликационной вилке. РНК-праймаза синтезирует РНК-затравки. ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи. ДНК-лигаза сшивает фрагменты после удаления РНК-затравки. Ключевыми событиями клеточного цикла как прокариот, так и эукариот являются репликация ДНК и деление клетки. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления. В подавляющем большинстве случаев прокариотические клетки делятся с образованием двух одинаковых по размеру и генетическому материалу дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. Раздел 2 Клеточный уровень организации живого 1. Клетка – элементарная открытая биологическая система. Типы клеточной организации. Основные положения клеточной теории. Клетка - основная структурная, функциональная и генетическая единица организации живого, элементарная живая система. Клетка может существовать как отдельный организм (бактерии, простейшие, некоторые водоросли и грибы) или в составе тканей многоклеточных животных, растений, грибов. Термин «клетка» был предложен английским исследователем Робертом Гуком в 1665 г. Впервые используя микроскоп для изучения срезов пробки, он заметил множество мелких образований, похожих на ячейки пчелиных сот. Роберт Гук дал им название ячейки или клетки. Работы Р. Гука вызвали интерес к дальнейшим микроскопическим исследованиям организмов. Возможности светового микроскопа в XVII-XVIII веках были ограничены. Накопление материала о клеточном строении растений и животных, о структуре самих клеток шло медленно. Только в тридцатых годах XIX века были сделаны фундаментальные обобщения о клеточной организации живого. Клеточная теория Основные положения клеточной теории сформулированы ботаником Матиасом Шлейденом (1838 г.) и зоологом-физиологом Теодором Шванном (1839 г.): • все организмы состоят из одинаковых структурных единиц - клеток; • клетки растений и животных сходны по строению, образуются и растут по одним и тем же законам. В 1858 г. немецкий ученый Рудольф Вирхов обосновал принцип преемственности клеток путем деления. Он писал: «Всякая клетка происходит из другой клетки ...», т.е. дал понять, откуда появляется клетка. Это утверждение стало третьим положением клеточной теории. Изучение клетки с помощью новейших физических и химических методов исследования позволили сформулировать основные положения современной клеточной теории: • все живые организмы состоят из клеток. Клетка — единица строения, функционирования, размножения и индивидуального развития живых организмов. Вне клетки нет жизни. • клетки всех организмов сходны между собой по строению и химическому составу; • на современном этапе развития живого клетки не могут образовываться из неклеточного вещества. Они появляются только из ранее существовавших клеток путем деления; • клеточное строение всех ныне живущих организмов - свидетельство един- ства происхождения. 2. Строение плазмолеммы: надмембранный, мембранный и суюмембранный комплексы. Медицинские аспекты функций плазмолеммы. Плазмолемма (внешняя клеточная мембрана, цитолемма, плазматическая мембрана)занимает в клетке пограничное положение и играет роль полупроницаемого селективного барьера, который, с одной стороны, отделяет цитоплазму от окружающей клетку среды, а с другой - обеспечивает ее связь с этой средой. Структура плазмолеммы.Плазмолемма - самая толстая из клеточных мембран (7.5-11 нм); под электронным микроскопом она, как и другие клеточные мембраны, имеет вид трехслойной структуры,представленной двумя электронно-плотными слоями, которые разделены светлым слоем. Ее молекулярное строение описывается жидкостно-мо-заичной моделью,согласно которой она состоит из липидного (фосфо-липидного) бислоя,в который погружены и с которым связаны молекулы белков. Липидныйбислойпредставлен преимущественно молекулами фосфатидилхолина (лецитина)и фосфатидилэтаноламина (цефалина),состоящими изгидрофильной (полярной) головкии гидрофобного (неполярного) хвоста.В состав большинства мембран входит также холестерин (холестерол).В мембране гидрофобные цепи обращены внутрь бислоя, а гидрофильные головки - кнаружи (рис.3-2). Состав липидов каждой из половин бислоя неидентичен. Липиды обеспечивают основные физико-химические свойства мембран, в частности, их текучестьпри температуре тела. Некоторые липиды (гликолипиды)связаны с олигосахаридными цепями, которые выступают за пределы наружной поверхности плазмолеммы, придавая ей асимметричность. Электронно-плотные слои соответствуют расположению гидрофильных участков липидных молекул. Мембранные белкисоставляют более 50% массы мембраны и удерживаются в липидном бислое за счет гидрофобных взаимодействий с молекулами липидов. Они обеспечивают специфические свойства мембраны(типы белков и их содержание в мембране отражают ее функцию) и играют различную биологическую роль (переносчиков, ферментов, рецепторов и структурных молекул).По своему расположению относительно липидного бислоя мембранные белки разделяются на две основные группы - интегральные и периферические. Периферические белкинепрочно связаны с поверхностью мембраны и обычно находятся вне липидного бислоя. Интегральные белкилибо полностью (собственно интегральные белки), либо частично (полуинтегральные белки)погружены в липидный бислой; часть белков целиком пронизывает всю мембрану (трансмембранные белки).Интегральные белки плазмолеммы хорошо выявляются при использовании метода замораживания-скалывания.При этом плоскость скола обычно проходит через гидрофобную середину бислоя, разделяя его на два листка - наружный и внутренний (см. рис. 3-2). Интегральные белки имеют вид округлых внутримембранных частиц,большая часть которых связана с Р'-поверхностью(от англ, protoplasmic) - протоплазматической, т.е. ближайшей к цитоплазме поверхности скола (наружной поверхности внутреннего листка), меньшая - на Е |