ихтиопаталогия. Дана характеристика рыб как переносчиков возбудителей болезней человека и животных. Для студентов вузов по специальности Водные биоресурсы и аквакультура
 Скачать 10.48 Mb. Скачать 10.48 Mb.
|

|
1.3.4. НЕКРОЗ Некроз — омертвение отдельных клеток или их групп, участков тканей и органов, наступающее при жизни организма. Физиологическое отмирание клеток и тканей происходит в результате старения и у здоровых рыб. Некроз же обусловлен повреждением клеток при действии на них различных патологических факторов. Факторы, вызывающие некроз, могут быть физическими (различные механические воздействия, травмы, резкие изменения условий обитания, различные химические вещества и др.) и биологическими (бактерии, вирусы, грибы, животные-паразиты). Например, некроз апикальных концов жаберных лепестков при бранхиомикозе, токсикозе, сангвиниколезе, дактилогирозе. Возникновение некроза во многом определяется характером тканей и органов, подвергающихся патологическим воздействиям, а также зависит от общего состояния организма. Так, быстрее и легче подвергаются некрозу эпителий, мышечная и жаберная ткани рыб. Наиболее подвержены некрозу истощенные рыбы, содержавшиеся в плохих условиях, или особи с нарушенным кровообращением, которое бывает при бранхиомикозе и других жаберных заболеваниях. Омертвению ткани предшествует более или менее длительный период некробиоза, характеризующийся развитием в тканях дистрофических процессов. Если в этот период устранить действие повреждающего фактора, то можно предотвратить наступление некроза. Гистологически основным признаком некроза тканей является исчезновение в их клетках ядер, что связано с растворением ядра (кариолизисом). Иногда ядро сморщивается, уплотняется (карио-пикноз), в дальнейшем распадается на отдельные глыбки и затем полностью исчезает (кариорексис). Соответственно изменяется Протоплазма клеток: вакуолизируется, постепенно распадается, превращаясь в мелкозернистую массу. При некрозе функция органа или его участка полностью прекращается. От места возникновения некроза и его распространения зависит состояние рыбы. К тяжелым последствиям приводят сильный некроз жабр при различных жаберных заболеваниях, некроз почек — при вирусной геморрагической септицемии форели, некроз стенок плавательного пузыря — при острой форме воспаления и др. Нередко на месте некроза развивается вторичная микрофлора, что отягчает патологический процесс. В некоторых случаях вокруг очага некроза разрастается соединительная ткань, образуется капсула, в которую заключен очаг некроза (например, при поражении печени форели триенофорусом). Позднее некротические массы уплотняются, в них откладываются известковые соли. 1.4. ЗАЩИТНЫЕ РЕАКЦИИ ОРГАНИЗМА Под термином «защитные реакции организма» скрываются два понятия: приспособительные функции и собственно защитные реакции. Первое, более широкое понятие определяет степень приспособленности (адаптивности) организма как биологической системы вида к среде, второе — относится к способностям организма защищать себя от негативного воздействия среды. Собственно защитные реакции организма включают в себя систему иммунитета, воспаление, гипертрофию, регенерацию и инкапсуляцию. 1.4.1. ИММУНИТЕТ Иммунитет является одним из основных защитных свойств организма. Он направлен против чужеродных веществ и организмов (антигенов), а также против собственных физиологически неполноценных элементов. Различают две основные формы иммунитета: врожденный и приобретенный. Врожденный иммунитет определяется генетически обусловленными морфологическими и биохимическими особенностями того или иного вида животных. Он проявляется в видовой, возрастной и индивидуальной устойчивости рыб к возбудителям заболеваний. Напряженность его значительно выше приобретенного, но она не является статической величиной и претерпевает различные колебания. Приобретенный иммунитет подразделяется на активный и пассивный. Активный иммунитет формируется после повторных воздействий на организм того или иного антигенного раздражения, отдаленных друг от друга различными интервалами времени, пассивный — с помощью введения готовых антител. В основе врожденной и приобретенной устойчивости лежит механизм взаимодействия многих гуморальных и клеточных факторов иммунитета, направленных на сохранение постоянства вНутренней среды организма на всех этапах индивидуального развития. Эта сложная система защитных факторов включается под влиянием антигенного раздражителя и направлена на его разрушение и выведение из организма. Прежде всего, патогену необходимо преодолеть физические и химические барьеры, составляющие первую линию зашиты против инвазии патогена. Особая роль отводится эпителиальным покровам кожи, жабр и пищеварительного тракта, которые в норме наряду с осморегуляцией играют важную роль в качестве механических средств противомикробной и противовирусной защиты. Не менее важной функцией эпителиальных слоев является продукция слизи, вырабатываемой слизистыми клетками. Слизь механически препятствует колонизации микроорганизмов на эпителиальных поверхностях, о чем свидетельствует усиление ее продукции в стрессовых условиях и под воздействием возбудителей инфекций. Кроме того, она имеет сложный биохимический состав, благодаря которому приобретает бактериолитические, антивирусные, противогрибковые и антипаразитарные свойства. Подобные характеристики слизи объясняются присутствием в ней ряда гуморальных факторов иммунитета: лизоцима, С-реактивного белка, иммуноглобулинов и комплемента. Все механизмы первой линии защиты неспецифичны и не обеспечивают развитие памяти к антигену в иммунной системе хозяина. Лизоцим — низкомолекулярный белок, обладающий выраженными ферментативными свойствами, молекула которого характеризуется высоким содержанием основных и дикарбоновых аминокислот. Лизоцим присутствует в сыворотке и слизи рыб, но в основном он ассоциируется с лейкоцитами (моноцитами, нейт-рофилами, макрофагами) и обогащенными данными группами клеток тканями. Лизоцим обладает антибактериальными свойствами, в основном относительно грамположительных бактерий, вызывая лизис микроорганизмов путем гидролиза нерастворимых полисахаридов клеточной оболочки или опсонизации патогена. С-реактивный белок (СРБ) выделяется как из крови, так и из слизи рыб. Биологические свойства данного протеина до конца не изучены. Тем не менее, имеются данные о том, что СРБ участвует в фагоцитозе, опсонизируя антигены; взаимодействует с комплиментарной системой, выступая в роли ее активатора; способствует лизису бактерий; повышает подвижность лейкоцитов и, что особенно важно, выступает в качестве «скрепляющего» компонента, агглютинируя патогены и препятствуя распространению инфекции. Процессы, обеспечивающие вторую линию защиты, активизируются только при внедрении патогена. Подобные механизмы функционируют на низком уровне специфичности и способны обеспечить развитие кратковременной памяти к антигену. Кровь, лимфа и ткани рыб содержат множество различных веществ, обладающих выраженным противомикробным действием (интерферон, хитиназа, С-реактивный белок и др.). Они различаются по химической природе и структуре и характеризуются определенной избирательностью. Комплекс данных веществ создает мощный гуморальный барьер. Так, например, бактерицидная активность сыворотки крови обеспечивается комплексным действием пропердина, комплемента, лизоцима и 3-лизина. Интерферон — низкомолекулярный клеточный белок, который синтезируется под влиянием различных индукторов, получивших название интерфероногенов. Основная функция интерферона в иммунной системе костистых рыб — противовирусная. Обладая ярко выраженным ингибирующим влиянием на размножение вирусов в чувствительных тканях, он прерывает процесс трансляции РНК, нарушая тем самым синтез вирусного белка. Хитиназа — внеклеточный фермент, локализованный в лим-фоидных тканях рыб. У рыб-зоопланктонофагов большое количество хитиназы обнаружено в слизистой кишечника, поджелудочной железе и крови. Помимо основной, пищеварительной функции, хитиназа способна выполнять и защитную, предохраняя рыб от поражения хитинсодержащими грибами и от инвазии паразитами. Система комплемента является важнейшим фактором резистентности организма в условиях как врожденного, так и приобретенного иммунитета. Комплементом называют сложный комплекс белков (около 20), образующих каскадные системы, формирующие быстрый многократно усиленный ответ на первичный сигнал. При этом продукт одной реакции служит катализатором последующей. В иммунном ответе рыб система комплемента участвует в реакции антиген-антитело, определяя скорость, характер и выраженность иммунной агрессии; в фагоцитозе активизирует фагоциты и опсонизирует антигены; лизирует бактериальные и другие чужеродные клетки, встраиваясь в их поверхностную мембрану. В настоящее время у рыб описаны три компонента комплемент-комплекса: О, СЗ, С5 и один промежуточный комплекс ЕА С 1, 2, 4. Результатом деятельности факторов второй линии защиты рыб являются воспаление и фагоцитоз. Барьерная функция воспаления препятствует распространению возбудителя заболевания из одного участка организма в другой. При этом в его очаге происходят значительные физико-химические изменения, носящие защитный характер. Избирательно меняется проницаемость кровеносных сосудов, в результате чего из сосудистого русла в очаг воспаления происходит выход белков и лейкоцитов. Отложения фибрина препятствуют дальнейшему продвижению инфекционного агента и становятся дополнительным избирательным фильтром. Основной реакцией организма, определяющей защитную роль воспаления, является фагоцитоз. Фагоцитоз подразумевает под собой захват и переваривание микроорганизмов и других чужеродных антигенов с последующим освобождением организма от их конечных продуктов распада. Основную фагоцитарную функцию в организме рыб несут на себе макрофаги и в меньшей степени микрофаги (фагоциты) — нейтрофилы и моноциты (см. цв. вкл. табл. II). Прикрепление микроорганизма к поверхности фагоцита (адгезия) происходит путем примитивного механизма узнавания, основанного на взаимодействии углеводных остатков. Прикрепившаяся к поверхностной мембране фагоцита частица инициирует фазу поглощения, которая происходит за счет образования псевдоподий, с последующим полным заключением в вакуоль (фагосому). Фагоцитоз может быть завершенным, когда возбудитель переваривается в фагосоме, или незавершенным, когда возбудитель в ней только изолируется, но сохраняет свою жизнеспособность и даже размножается. Этот защитный механизм очень неустойчив, и при неблагоприятных условиях сохранившие жизнеспособность микроорганизмы могут вызывать обострение заболевания. Механизмы, составляющие третью линию защиты организма рыб, являются высокоспецифичными иммунными реакциями. В ответ на проникновение патогенов и чужеродных веществ (антигенов) в организме рыб вырабатываются антитела или антителопо-добные структуры (рецепторы). Антитела циркулируют в организме, а рецепторы находятся на поверхности так называемых сенсибилизированных клеток. Основной особенностью антител и рецепторов сенсибилизированных клеток является наличие в их молекуле участков (активных центров), точно соответствующих (специфичных) определенным участкам (детерминантам) в молекуле антигена. Третья линия защиты обеспечивается посредством лимфоидных клеток, индуцирующих возникновение иммунной памяти. В общих чертах активизация подобной защиты происходит следующим образом: 1. Воздействие антигена на организм стимулирует определенное число предетерминированных лимфоцитов, способных распознавать антиген посредством специфичных к нему рецепторов. 2. Под действием стимуляции происходит пролиферация клона лимфоцитов с дифференциацией дочерних клеток, которые в свою очередь несут на себе определенную функцию в зависимости от популяции. У рыб скопления лимфоцитов находятся в тимусе, головной и туловищной почках, селезенке и в стенках кишечника. При этом выделяют две основные их популяции: Т- (тимус-зависимые) и В-лимфоциты, чаще всего ассоциирующиеся у рыб с почками. Функции вышеуказанных популяций лимфоцитов весьма полно изучены у теплокровных и в меньшей степени у рыб: 1) В-лимфоциты являются продуцентами антител или иммуноглобулинов; 2) Т-лимфоциты, не продуцируя антитела, дифференцируются на несколько типов функциональных клеток, включающих в себя: цитотоксические «клетки-киллеры», непосредственно участвующие в лизисе чужеродных частиц; «лимфокин-проду-цирующие клетки», усиливающие неспецифическую активность макрофагов посредством продукции лимфокинов (интер-лейкинов); «Т-супрессоры», регулирующие продукцию антител и лимфокинов; «Т-хелперы», или «Т-помощники», взаимодействующие с В-лимфоцитами и усиливающие их ответ на введение антигена; 3) продукция антител требует присутствия третьей клеточной популяции, «перерабатывающей» и «подающей» антиген, которая у рыб представлена макрофагами и моноцитами. Эта фаза иммунного ответа протекает при прямом участии молекул главного комплекса гистосовместимости, играющего ключевую роль в распознавании «чужого» материала. Пролиферация и дифференцировка иммунокомпетентных клеток (лимфоцитов, цитотоксических клеток, моноцитов, макрофагов, гранулоцитов) осуществляется у рыб главным образом в почке, селезенке и тимусе. Воздействие антигена приводит к образованию так называемых Т- и В-клеток памяти, которые при вторичном инфицировании обеспечивают более быструю продукцию антител и в более высоких титрах. По своему происхождению антитела делятся на естественные (нормальные) и приобретенные. Естественные антитела образуются в ответ на малые количества антигенного раздражителя, т. е. при субклинической инфекции. К приобретенным антителам относятся все иммуноглобулины, появляющиеся в организме рыб в ответ на введение различных по своей структуре и природе антигенов или в результате перенесения того или иного инфекционного заболевания. Независимо от своего происхождения антитела относятся к гликопротеидам и содержат различные количества олигосахаридов разного состава и строения. Антитела, образующиеся в организме животных, различаются между собой по первичной структуре полипептидных цепей, физико-химическим свойствам и антигенной структуре. Для костистых рыб описан единственный класс иммуноглобулинов — тетрамерный макроглобулин, подобный иммуноглобулинам класса М (IgM) млекопитающих. В зависимости от вида рыб его молекулярный вес колеблется от 600 до 800 кД. Некоторые рыбы имеют мономерные или диамерные формы иммуноглобулинов, которые часто обнаруживаются в эпидермальной слизи, кишечнике и желчи. Основными функциями антител являются взаимодействие с антигеном и его инактивация посредством нейтрализации (вирусы, токсины), агглютинации (бактерии) и преципитации (растворимые антигены), что в свою очередь усиливает реакцию фагоцитоза за счет более быстрого распознавания чужеродных частиц и активирует систему комплемента, приводя к лизису чужеродные клетки. Интенсивность антителообразования определяется видовой реактивностью организма рыб, температурными условиями среды их обитания, интенсивностью и длительностью антигенного раздражения и другими факторами. Иммунологические методы в аквакультуре используют для диагностики и профилактики болезней рыб. В основе диагностики лежат специфические серологические реакции, позволяющие выявлять антигены и антитела к определенным возбудителям и определять природу возбудителя болезни. В результате взаимодействия специфических антител и антигена образуется комплекс антиген-антитело, или иммунный комплекс (рис. 1). Методы выявления специфических антител направлены на их взаимодействие с соответствующим антигеном, который называется диагностическим антигеном. Если у обследованной рыбы есть специфические антитела, то они соединятся с диагностическим антигеном и образуют иммунный комплекс. Образование иммунного комплекса в диагностической реакции сопровождается выпадением осадка, помутнением или другими явлениями и свидетельствует о том, что ранее рыба была носителем или контактировала с возбудителем. Чтобы обнаружить антиген (т.е. возбудитель), нужно иметь диагностические антитела, которые получают путем иммунизации (т. е. специальным заражением данным возбудителем) животных (рыб, кроликов, мышей и т. д.) или культивирования культур гибридных клеток — гибридом. Рис. 1. Схема определения антител (а) и антигенов (б) 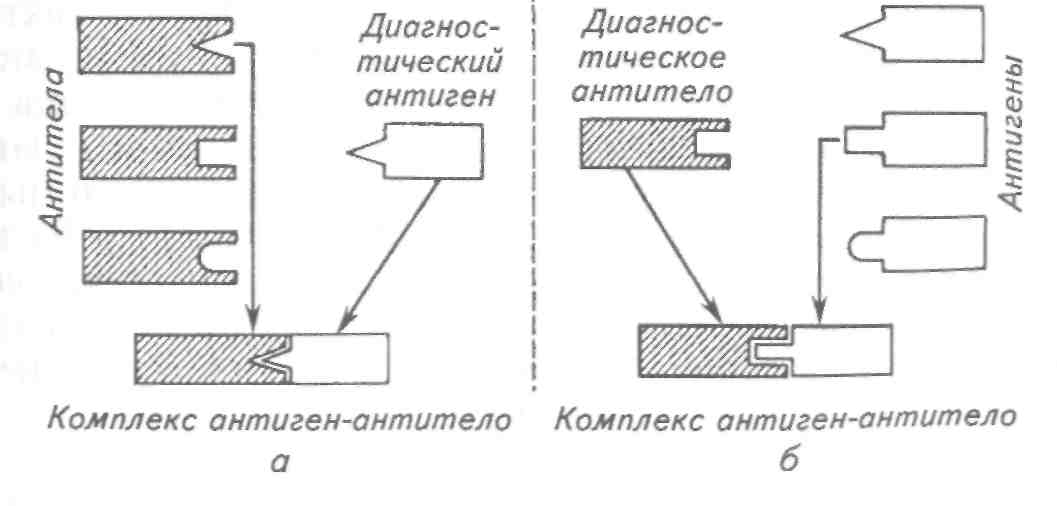 У иммунизированных животных в период наибольшего содержания антител берут кровь и получают из нее сыворотку, которую называют гипериммунной диагностической сывороткой. Диагностические моноклональные антитела получают путем культивирования культур гибридных клеток — гибридом. Если у обследуемой рыбы имеется специфический антиген, то он связывается с диагностическими антителами и образует иммунный комплекс. За рубежом выпускают целый ряд диагностических наборов (диагностикумов) для особо опасных вирусных и бактериальных инфекций. В последнее время активно разрабатываются диагностикумы для обнаружения генетического материала патогенов рыб, основанные на методах гибридизации нукло-новых клеток и полимерозной цепной реакции. Однако следует отметить, что эти методы являются творением генной инженерии и при всей их высокой специфичности и чувствительности они не относятся к методам иммунологической диагностики. В нашей стране для промышленного использования рекомендован диагностикум на вибриоз и проводятся производственные испытания диагностикума на весеннюю виремию карпа. Успешное применение иммунологических исследований нашло отражение в иммунопрофилактике. Для повышения неспецифической резистентности в организм рыб вводят иммуностимуляторы, которые повышают активность иммунологических реакций, в том числе клеточных и гуморальных факторов иммунитета. В мировой аквакультуре неспецифическая иммунопрофилактика нашла широкое применение. Промышленностью освоено несколько таких препаратов, которые задают рыбе с кормом (левамизол, глюканы, хитозан и др.). Сюда же относится ряд иммуностимулирующих витаминов, доказавших свою эффективность на рыбах (С, Е и А). Вакцинопрофилактика основана на иммунологической специфичности и памяти. Благодаря клеткам памяти иммунная система способна гораздо сильнее отвечать на повторную встречу с данным антигеном. При создании вакцины возбудитель заболевания изменяют таким образом, чтобы он стал безвредным (инактивиру-ют, аттенуируют), но не потерял своей антигенности, либо используют изолированные антигены возбудителя. С конца 1980-х годов начало активно развиваться новое направление в вакцинологии рыб — рекомбинантные генно-инженерные вакцины. В ответ на введение вакцины в организме рыбы вырабатываются специфические антитела. В случае последующего естественного заражения этим возбудителем он рестимулирует клетки памяти, вызывая вторичное, более быстрое и сильное образование антител, которые нейтрализуют возбудителя, образуя иммунный комплекс. Для профилактики особо опасных болезней рыб в мире разработано около 20 вакцинных препаратов, часть из которых рекомендована для лабораторного, а часть — для коммерческого использования. В производственных условиях применяют вакцины против вибриоза, холодноводного вибриоза, фурункулеза, иерси-ниоза, а также против инфекционного некроза поджелудочной железы и вирусной геморрагической септицемии. В нашей стране хорошие результаты получены при производственных испытаниях биохимической пртивоаэромоидной вакцины ВЮС-2. Ведутся работы по созданию рекомбинантной генно-инженерной вакцины против весенней виремии карпа. 1.4.2. ВОСПАЛЕНИЕ Воспаление — это местная защитная реакция в ткани или органе, возникающая в ответ на воздействие повреждающих факторов. Оно относится к деятельности второй линии защиты и присуще всем организмам, на какой бы ступени эволюционной лестницы они не стояли. И. И. Мечников (1917) считал, что это древнейшая «спасительная» реакция живых организмов. Неспособность организма отреагировать на патогенное воздействие воспалительной реакцией грозит гибелью. Воспаление состоит из трех патологических процессов: повреждения тканей (альтерация), сосудистых изменений (экссудация) и размножения клеточных элементов (пролиферация). В ходе каждого из них в поврежденной ткани проходят важные физиолого-биохимические процессы. Так, в процессе альтерации (деструктивный процесс) высвобождается значительное количество химически активных веществ (медиаторов), изменяющих рН среды, стимулирующих деятельность фагоцитов, обеспечивающих непрерывность кровотока, и развивается гиперемия. Процесс экссудации сопровождается нарушением проницаемости кровеносных сосудов, выходом из них жидкой фракции плазмы крови и лейкоцитов. В очаге воспаления скапливается значительное количество жидкости и фагоцитирующих клеток. Сюда привлекаются не только лейкоциты (микрофаги), но и макрофаги. В результате экссудации повышается вязкость крови и кровоток замедляется. Нарушается распределение форменных элементов в кровеносных сосудах. Если при нормальном кровотоке форменные элементы располагаются по центру сосуда, то после выпотевания плазмы и соответственно увеличения вязкости крови они перемещаются к периферии сосуда. Лейкоциты прилипают к внутренней стенке сосуда и затем выходят за его пределы. Вышедшая из кровеносных сосудов жидкость (воспалительный выпот), скапливающаяся в каких-либо полостях или тканевых щелях, называйся экссудатом, а если она пропитывает ткань — инфильтратом. Экссудат содержит значительное количество белка. Процесс пролиферации характеризуется активным размножением клеток в основном соединительной ткани. Причиной размножения клеток считают обилие в очаге воспаления питательных веществ и биологических стимуляторов, скапливающихся здесь в результате альтеративных и экссудативных процессов. Кроме того, благодаря воспалительной гиперемии в очаге воспаления усиливается обмен веществ, что также стимулирует размножение клеток. Размножающиеся клетки соединительной ткани выполняют роль фагоцитов (защитная функция), а также создают клеточный барьер между очагом воспаления и здоровыми тканями. Описанные процессы воспалительной реакции протекают не последовательно друг за другом. Так, пролиферативные явления могут проявляться в самом начале с момента повреждения тканей, т.е. одновременно с альтерацией. Экссудативный процесс также может проявиться в начале альтерации или затянуться до затухания пролиферативного процесса. Только морфологические, патофизиологические и биохимические исследования позволяют раскрыть сложные механизмы, играющие роль в развитии воспаления. Воспаление, хотя и местный процесс, но он оказывает влияние на организм в целом. Развиваются изменения, носящие общий характер: меняется состав крови, кровоснабжение, обмен веществ и т. д. У рыб воспалительный процесс изучен далеко не достаточно. Ему во многом присущи внешние признаки воспаления (покраснение, припухлость, расстройство функций). Причинами воспаления являются любые воздействия на орган или ткань рыбы, превышающие их устойчивость. Возможны физические причины (механические, термические), химические (воздействие токсических веществ, нарушения газового и химического режимов в водоеме), биологические (воздействие инфекционных и инвазионных патогенных агентов). Перечисленные причины относятся к группе внешних (экзогенных) факторов. Существует группа внутренних (эндогенных) факторов, но эти причины возникновения воспалительной реакции у рыб не изучены. Воспаление проявляется в двух формах: острой и хронической. Форма течения воспалительной реакции зависит от причины, ее вызвавшей, и физиологического состояния организма. Острое течение характеризуется преобладанием альтеративных и экссудативных явлений, протекает быстро. Хроническое течение обычно растянуто во времени, и в нем преобладают атрофические и пролиферативные изменения. Воспаление может заканчиваться полным восстановлением ткани (органа), или исход его может быть неполным. В этом случае остается патологическое состояние ткани (органа) в виде про-лиферативных разрастаний (соединительнотканные спайки, сращения), или атрофии. В зависимости от того, какой из трех патологических процессов преобладает в конкретной воспалительной реакции, различают три основные его формы: альтеративное, экссудативное и проли-феративное. Альтеративное воспаление часто возникает при инфекционных процессах или действии на организм токсических веществ. В нем преобладают дегенеративно-некротические процессы. Хорошо выражены гиперемия, слущивание эпителия со слизистых оболочек. Слабо проявляются эмиграция из сосудов лейкоцитов и пролиферация. Чаще всего эта форма воспаления протекает в острой форме. Однако при хроническом течении возможно замещение клеточных элементов ткани органа соединительной тканью. Особенно четко альтеративные изменения бывают выражены в паренхиматозных органах — печени, почках, селезенке. Экссудативное воспаление характеризуется нарушением проницаемости стенок кровеносных сосудов и хорошо выраженным выходом из них составных элементов крови. Этот вид воспаления наблюдается в органах с хорошо развитой кровеносной системой (слизистые оболочки, жабры, плавательный пузырь). В зависимости от составных частей экссудата воспаление этой формы может быть серозным, фибринозным, гнойным и геморрагическим. Серозное воспаление отличается выделением экссудата, содержащего небольшое количество белка (3—5 %) и клеточных элементов крови. При этом воспалении развивается заметный отек. Серозному воспалению в большой степени подвержены слизистые оболочки. Причинами могут стать механические повреждения, температурный фактор, токсины, инфекционные агенты. Фибринозное воспаление подразумевает содержание в экссудате большого количества белка — фибриногена, который свертывается и откладывается на серозных и слизистых оболочках, между тканевыми элементами. Пленка свернувшегося фибриногена может быть очень плотной, и в результате в пораженных слизистых оболочках развивается некроз. Гнойное воспаление возникает в поверхностных слоях тканей и в их толще. Оно характеризуется большим скоплением в очаге воспаления лейкоцитов в состоянии распада и гнойной сыворотки, которая содержит много белка и неспособна к свертыванию. Все это образует гной — мутную густую жидкость желтовато-зеленого цвета. При гнойном воспалении возможно возникновение абсцесса — полости в ткани, заполненной гноем, или флегмоны — пропитывание тканей гноем. Причиной гнойного воспаления чаще всего является воздействие гноеродных микроорганизмов, например при фурункулезе лососевых. Геморрагическое воспаление — это наиболее тяжелая форма экссудативного воспаления, когда в экссудате содержится большое количество эритроцитов, что придает ему красноватый оттенок. Причиной этого воспаления, как правило, являются патогенные микроорганизмы. При этом проницаемость кровеносных сосудов настолько повышается, что через них выходят не только Плазма и лейкоциты, но и эритроциты. У рыб геморрагическое воспаление наблюдается у лососевых при некоторых вирусных заболеваниях, у карпа при эритродерматите. Пролиферативное (продуктивное) воспаление наблюдается при преобладании в очаге воспаления процессов клеточного размножения и в результате разрастания местной ткани. Разрастающаяся ткань богата кровеносными сосудами, содержит большое количество гистиоцитов, плазматических эпителиальных клеток. Часто присутствуют гигантские клетки с множеством ядер. Здесь же скапливаются различные формы лейкоцитов. Со временем разрушенная паренхима органа замещается соединительной тканью. Пролиферативное воспаление наблюдается при проникновении в ткани различных паразитических животных. При этом сначала развиваются альтеративные и экссудативные процессы, сменяющиеся пролиферацией, которая носит характер отграничивающей реакции и завершается образованием капсулы, окружающей паразита. Подобное явление наблюдается при паразитировании плероцерко-идов цестод, метацеркариев трематод в мускулатуре или во внутренних органах некоторых рыб. Со временем в этих соединительнотканных капсулах проходят дистрофические изменения: они могут обызвествляться (процесс петрификации), а находящиеся внутри них паразиты погибают и постепенно рассасываются. 1.4.3. ГИПЕРТРОФИЯ Гипертрофией называют увеличение объема ткани или органа. Различают собственно гипертрофию, при которой увеличение объема ткани или органа происходит за счет увеличения объема его клеток и тканевых элементов, и гиперплазию, когда увеличение органа или ткани происходит вследствие размножения и увеличения количества его клеток и тканевых элементов. Гипертрофия может быть физиологической и патологической. При физиологической гипертрофии увеличение объема органа происходит в результате усиления его функции под влиянием естественных причин (например, увеличение размера гонад у рыб перед икрометанием). Особенностью физиологической гипертрофии является ее обратимость. После прекращения действия причин, ее вызывающих, орган или ткань возвращается в нормальное состояние. Патологическая гипертрофия развивается под влиянием каких-либо чрезмерных для организма факторов. В этих случаях в тканях развивается временное усиление обмена веществ (когда организм пытается справиться с непривычным раздражителем), а затем гипертрофический процесс переходит в атрофию или дистрофию, нарушается нейрогуморальная регуляция и процесс становится необратимым. У рыб патологическая гипертрофия наблюдается при некоторых вирусных заболеваниях (лимфоцистис, стоматопа-пиллома угрей) и при паразитировании внутриклеточных простейших (микроспоридии). 1.4.4. РЕГЕНЕРАЦИЯ Регенерацией называют восстановление утраченных или поврежденных частей тела. Это защитная (приспособительная) реакция выработана в процессе эволюционного развития организмов. У позвоночных возможно восстановление только тканей. Причем быстрее восстанавливаются ткани, которые часто повреждаются (эпидермис, слизистые оболочки), а более защищенные ткани (нервные клетки, хрящевые ткани, сердечная мышца) обладают наименьшей способностью к регенерации. У рыб регенерация проявляется в восстановлении разрушенных жаберных лепестков, жаберных крышек, плавников, кожных покровов и др. Регенерация может быть физиологической и восстановительной (репаративной). Физиологическая регенерация — это естественный процесс замещения новообразованными тканевыми элементами старых, утраченных в результате физиологического некроза. Этот процесс совершается в организме постоянно (отмирание клеток эпидермиса, клеток красной и белой крови и т. д.). В организме существуют специальные тканевые системы и органы (центры регенерации), в которых воспроизводятся новые клетки взамен подвергшихся естественному отмиранию. Особенности физиологической регенерации заключаются в том, что вновь образованные клеточные элементы полностью замещают утраченные не только в количественном, но и в качественном отношении. Как правило, они более соответствуют состоянию организма (например, возрасту) и особенностям внешних условий (например, сезону года, изменениям гидрологических условий в водоеме). Восстановительная (репаративная) регенерация — это замещение клеточных элементов и тканей, разрушенных в результате воздействия вредных факторов. Она может быть полной и неполной. При полной регенерации поврежденная ткань замещается идентичной, полностью соответствуя утраченной. Это происходит при незначительном повреждении, сохранении нервного и сосудистого аппаратов. При неполной регенерации происходит замещение дефекта тканью, отличающейся от утраченной (чаще всего волокнистой соединительной тканью), или с выраженными отклонениями от нормального строения. Данный вид регенерации наблюдается при обширных повреждениях, сопровождающихся нарушением нервных и сосудистых элементов. Скорость восстановительного Процесса различна и зависит от свойств поврежденной ткани и степени ее разрушения. Некоторые ткани, например, нервная, хРящевая, регенерируют медленно, а эпителиальная, железистая и Другие восстанавливаются довольно быстро. На процесс регенерации оказывают существенное воздействие условия окружающей среды, наличие полноценного питания, физиологический статус, возраст. У молодых регенерация тканей происходит быстрее, чем у Рыб старших возрастов. 1.4.5. ИНКАПСУЛЯЦИЯ Инкапсуляцией называется обрастание тканью патологического очага и образование вокруг него капсулы. Капсула может окружать доброкачественную опухоль, очаг воспаления, инородное тело, попавшее в ткань, или паразита и ослаблять их воздействие на организм. Образование капсул связано в основном с пролифе-ративными процессами воспаления, реже с патологической гипертрофией и избыточной регенерацией. Развитие процесса инкапсуляции является не только ответом на механическое раздражение тканей, но и специфической защитной реакцией организма на ограничение и возможное подавление вредоносного воздействия патологического очага (или патогенного агента). Обрастание паразитов тканью хозяина, т.е. образование вокруг него капсулы, наблюдается у рыб весьма часто. Различают капсулы эпителиальные, соединительнотканные и смешанного происхождения. Эпителиальные капсулы выделяются как кожными покровами, так и кишечным эпителием. В качестве примера такой капсулы можно привести разрастание кожной ткани вокруг личинок двустворчатых моллюсков — глохидий, паразитирующих на плавниках и в жабрах различных пресноводных рыб. Глохидий с помощью своих створок зажимают участок межплавниковой ткани или жабр рыбы. В результате повреждения ткани начинается ее разрастание вокруг глохидий и образуется эпителиальная капсула. При поражении ихтиофтириусом также наблюдается разрастание эпителия, лежащего над паразитом, с образованием небольшого бугорка. При сильном заражении этим паразитом на жабрах начинается разрастание респираторных складок, что иногда приводит даже к срастанию образовавшихся выростов. В результате этого процесса паразит как бы оттесняется от наиболее важных участков ткани и погибает. Разрастание эпителиальной ткани может приобретать форму выростов. Характерные выросты образуются у мальков карпа под влиянием моногенеи Dactylogyrus vastator. Паразит прикрепляется к вершине жаберного лепестка, который начинает бурно расти. В результате возникают прозрачные длинные тяжи, на которых паразит не может удержаться и падает, после чего отпадают и выросты. В результате малек от него освобождается. Часто встречаются у рыб соединительнотканные капсулы. Они образуются под влиянием некоторых бактерий, грибов (ихтиофо-нуса), простейших (миксоспоридий и микроспоридий), плероцер-коидов цестод, матацеркариев трематод, личинок нематод и скребней, паразитирующих в полости тела, в стенках кишечника и во внутренних органах. Обычно капсулы приобретают округлую форму. Таковы белые паренхиматозные капсулы, окружающие плероцеркоидов цестод рода Triaenophorus и Diphyllobothrium, достигающие величины горошины, иногда и больше, видимые невооруженным глазом. Капсулы, образующиеся вокруг личинок нематод, вытянуты в длину, приобретают форму чехлика. Они бывают иногда совсем прозрачными и даже окрашенными в очень яркие цвета: оранжевый, зеленый, синий. У рыб наблюдают также капсулообразные, эпителиально-соединительнотканные разрастания. Например, они выявлены на жаберных лепестках белого амура и толстолобика, в местах прикрепления рачков Sinergasilus lieni, S. major. КОНТРОЛЬНЫЕ ВОПРОСЫ И ЗАДАНИЯ 1. Дать определение понятия болезнь. 2. Перечислить основные факторы, вызывающие заболевания у рыб. 3. Рассказать о принципах современной классификации болезней рыб. 4. Перечислить основные приемы диагностики болезней рыб. 5. Рассказать о нарушениях обмена веществ. 6. Охарактеризуйте основные патологические процессы. 7. Что собой представляет иммунитет? |