Физиология дыхания
 Скачать 1.59 Mb. Скачать 1.59 Mb.
|

|
3.1 Транспорт кислорода кровью. Кислород присутствует в крови в двух формах: растворенный в плазме и в воде, содержащейся в эритроцитах; а также в виде обратимой связи с молекулами гемоглобина в эритроцитах. Растворимость кислорода в жидкости при 37°C очень низкая и составляет 0,225 мл/л-1·кПа-1 (3 мл O2в 1 л плазмы), что недостаточно для жизнедеятельности организма в целом, поскольку в покое в организме человека в минуту потребляется примерно 250 мл кислорода. Проблема транспорта кислорода была более эффективно решена за счет обратимой химической реакции с гемоглобином эритроцитов. Гемоглобин - это хромопротеид, представляющий собой тетрамер. Каждый мономер состоит из гема и глобина. Гем - это порфириновое соединение, связанное с одним атомом железа. Глобин - это полипептид, представляющий собой α-цепь (141 аминокислота) и β-цепь (146 аминокислот). Гомология между α-и β-цепью достаточно высока, что дает возможность цепям иметь одинаковые конформации. Таким образом, комплекс гемоглобина имеет форму [α(Гем)]2[β (Гем)]2 и может связывать 4 молекулы О2. При обратимой реакции присоединенная к гему молекула О2 связывается с атомом железа, которое находится во второй степени окисления (т.е. это двухвалентное железо Fe2+). Соединение, образовавшееся в результате связывания называется оксигемоглобин (HbO2), тогда как гемоглобин без О2 называется дезоксигемоглобин (Hb). Это присоединение О2, которое проходит без изменения степени окисления (оксидации) железа, называется оксигенацией (это не оксидация или окисление), отщепление О2 - дезоксигенация. В 96-98% случаев гемоглобин взрослых представляет собой форму HbA1, глобин которого состоит из 2 α-субъединиц и 2 β-субъединиц. В отличие от глобина взрослых , глобин плода гемоглобина HbF состоит из двух α-цепей и двух γ-цепей. Оксигенированная кровь - светло-красная, тогда как дезоксигенированная - синевато-темнокрасная (синюшно-багровый цвет НЬ). Если абсолютная концентрация дезоксигенированного гемоглобина в капиллярной крови повышается более, чем на 50 г/л, то это приводит к посинению кожи и слизистых покровов (цианоз). Связывание кислорода гемоглобином. Наиболее важным фактором насыщения гемоглобина кислородом является pO2крови. Общее количество кислорода, переносимого гемоглобином в крови, зависит также содержания гемоглобина в крови. Чтобы лучше представить свойства реакции связывания О2 гемоглобином, принимают во внимание только концентрацию О2, связанного с гемоглобином, это значит, концентрацию оксигенированного гема в гемоглобине (HbO2). Ее можно соотнести к кислородной емкости крови, что значит к общей, способной к связыванию, концентрации гемоглобина, и таким образом, получить насыщение О2: 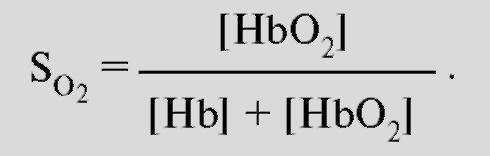 Экспериментально установлена количественная взаимосвязь между этими величинами, которая называется кривой диссоциации оксигемоглобина (рис.7). Кривая диссоциации HbO2 имеет крутой наклон между 10 и 60 мм рт.ст. pO2 относительно пологой части – между 70 и 100 мм рт. ст. Это обусловлено тем, что при pO2 60 мм рт. ст. содержание оксигемоглобина достигает порядка 90%., после чего дальнейшее увеличение pO2 вызывает незначительное связывание кислорода с гемоглобином. Количество кислорода, которое может связаться с гемоглобином эритроцитов при насыщении 100% его количества, называется кислородной емкостью гемоглобина. 1 г гемоглобина связывает 1,34мл кислорода. Поэтому в норме при концентрации гемоглобина 150г/л каждые 100мл крови могут переносить 20,0мл кислорода. Кислородная емкость крови отражает истинное количество кислорода в крови, как связанного с гемоглобином, так и физически растворенного в плазме. Поскольку в норме артериальная кровь насыщена кислородом на 97%, то в 100 мл крови содержится примерно 19,4мл кислорода. 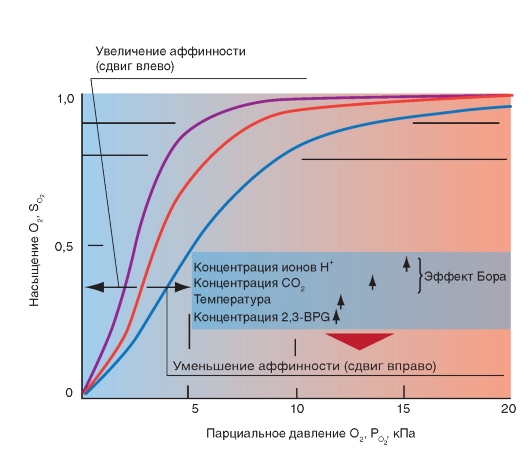 Рис.7. Кривая диссоциации оксигемоглобина. На кривой связывания О2 (сатурационная кривая) крови (рис.8) показана зависимость между насыщением гемоглобина О2 (SO2 ) от РО2 , так что кривая отражает кислородную емкость крови при разных значениях РО2 . Кривая имеет S-образную форму и парциальное давление полунасыщения (Р0,5) в 3,6 кПа. Далее при увеличении значения РО2 насыщение гемоглобина кислородом увеличивается от 75 до 98%. Кривая действительна для приведенных значений, характерных для нормальной артериальной крови. Для сравнения приведена кривая связывания кислорода миоглобином, имеющая форму гиперболы. Как видно из нижнего участка кривой, одинаковой с артериальной кровью , SO2 достигается при меньших значениях РО2 . Как видно из верхнего участка кривой связывания кислорода миоглобином, насыщение SO2 мало зависит от РО2. На связывание гемоглобином кислорода (аффинность, сродство) влияет ряд факторов. При этом, в первую очередь, изменяется положение кривой и форма кривой, но в меньшей степени. Отсюда можно описать уменьшение аффинности как сдвиг кривой связывания гемоглобином кислорода вправо (повышенный Р0.5), а увеличение аффинности, как сдвиг кривой связывания гемоглобином кислорода влево (пониженный Р0,5). Повышение температуры приводит к понижению аффинности, т.е. приводит к сдвигу кривой связывания вправо (рис. 7). Наоборот, охлаждение вызывает сдвиг кривой влево. У гомойотермных организмов значение этого влияния небольшое, хотя температура крови на периферии тела отчетливо может отклоняться от температуры всего организма. На связывание кислорода гемоглобином оказывает влияние 2,3-дифосфоглицерат. 2,3-дифосфоглицерат образуется в эритроцитах при гликолизе. 2,3-дифосфоглицерат обратимо связывается с гемоглобином, понижая его сродство к кислороду. Содержание в эритроцитах 2,3-дифосфоглицерата увеличивается при гипоксии, например, при пребывании человека в условиях высокогорья, что является одним из факторов адаптации. 2,3-дифосфоглицерат, понижая сродство гемоглобина к кислороду, способствует более легкой отдаче кислорода клеткам тканей и их нормальному кислородному снабжению в условиях недостаточного поступления в организм человека кислорода через легкие. . 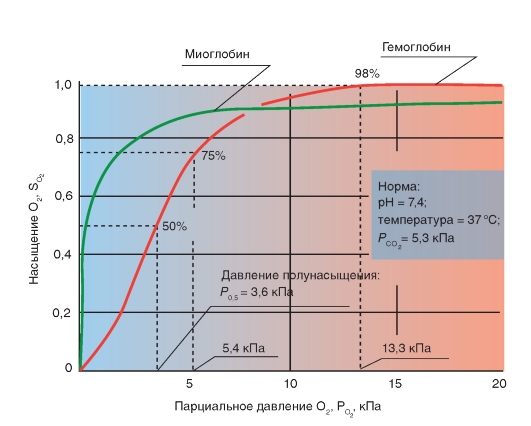 Рис. 8. Сатурационная кривая О2. Более важным является влияние концентрации ионов Н+ и РСО2. Повышение концентрации ионов Н+ (понижение рН) вызывает снижение аффинности гемоглобина к кислороду (сдвиг кривой вправо), понижение концентрации ионов Н+ вызывает повышение аффинности гемоглобина к кислороду. Эта зависимость, известная как эффект Бора, осуществляется путем аллостерического обменного влияния (термин «аллостерический» означает «связанный с другим центром») между местом связывания Н+ и местом связывания О2. Эффект Бора может быть связан также с изменением РСО2: повышение РСО2сдвигает кривую связывания О2 вправо, понижение - влево. Это влияние основывается, в первую очередь, на том, что повышение РСО2 ведет к понижению рН и наоборот. Таким образом, уменьшение аффинности означает, что при одинаковом РО2 гемоглобин способен связывать О2 в меньшей степени; кривая при этом сдвигается вправо. Причиной этого является повышение ряда факторов в эритроците (t°, рН, CO2 и т.п.). Уменьшение этих факторов влияет, наоборот, на повышение аффинности, т.е. сдвиг кривой влево. 3.2.Транспорт углекислого газа от тканей к легким. В покое у человека в процессе метаболизма образуется 200 мл СO2 в минуту. СO2 имеет растворимость в воде выше, чем кислород, и поэтому в большем количестве переносится плазмой крови и эритроцитами в физически растворенном состоянии (около 10%). Примерно 30% молекул СO2 , поступающих в кровь, обратимо связывается с аминогруппами гемоглобина, образуя карбаминогемоглобин (карбгемоглобин):СO2+Hb= HbCO2, поскольку восстановленный гемоглобин имеет высокое сродство к СO2. Остальные 60% молекул углекислого газа, поступающих в кровь, превращаются в бикарбонат и в такой форме СO2транспортируется к легким. CO2 из тканей диффундирует по градиенту концентрации внутрь эритроцитов. В эритроцитах реакция СO2 + Н2О=Н2СО3= НСО3- + Н+ осуществляется с обязательным участием фермента карбоангидразы, которая содержится в мембранах эритроцитов. Затем угольная кислота диссоциирует на ион водорода и ион бикарбоната. Большинство ионов бикарбоната по мере образования выводится через мембрану эритроцитов в плазму крови с помощью белка транспортера. При этом один ион бикарбоната обменивается на один ион хлора (хлорный сдвиг). Ионы H+ и CO2 связываются с молекулой гемоглобина, образуя карбаминовое соединение, в виде которого переносится часть CO2 к легочным капиллярам. Вторая фракция переноса CO2 от тканей к легким – это HCO3-. HCO3- поступают в плазму крови и связываются с ионами Na+ и участвуют в регуляции pH крови.CO2 практически не выводится из организма таким путем. Основными источниками выдыхаемого CO2 являются соединения CO2 и H+- ионов с гемоглобином эритроцитов, а также физически растворенная в плазме крови фракция CO2. В капиллярах легких в эритроцитах происходит диссоциация соединений CO2 с гемоглобином и H+- ионами (эффект Бора). Напряжение CO2 в венозной крови выше, чем в альвеолярном воздухе. Физически растворенный углекислый газ диффундирует через альвеолярную мембрану в просвет альвеолярного пространства, а затем выводится из легких с выдыхаемым воздухом. Выведение CO2 из венозной крови в альвеолы сдвигает реакцию CO2+ H2O=H2CO3=HCO3-+H+ влево с образованием H2O и CO2. CО2 диффундирует из крови в альвеолы. Карбаминовые соединения гемоглобина высвобождают CO2, и углекислый газ диффундирует в альвеолы, а затем выводится из легких во внешнюю среду. 3.3.Транспорт ионов водорода от тканей к легким. Другие виды гемоглобина. Восстановленный гемоглобин имеет более высокое сродство к протонам, чем оксигемоглобин, и связывает большинство протонов, которые образуются в вышеуказанной реакции. В результате только незначительное количество ионов водорода находится в крови в свободном состоянии. Это объясняет тот факт, что кислотность венозной крови незначительно ниже, чем в артериальной крови. В капиллярах легких – по мере образования оксигемоглобина реакция HbO2+ H+=HbH+ O2 протекает в обратном направлении с образованием оксигемоглобина и ионов водорода. Ионы водорода реагируют с ионами бикарбоната, что сдвигает реакцию влево с образованием воды и углекислого газа, который диффундирует в альвеолы и удаляется из организма с выдыхаемым воздухом. Гемоглобин может образовывать также соединения с угарным газом (окись углерода, монооксид углерода, CO)и взаимодействовать с сильными окислителями с изменением валентности железа в геме. Соединение гемоглобина с угарным газом называется карбоксигемоглобином (HbCO). Аффинность гемоглогбина к CO приблизительно в 300 раз больше, чем O2. , т.е.гемоглобин связывает CO более активно. В этом случае гемоглобин не может быть использован для транспорта кислорода. Соединение CO с железом гема ведет к увеличению аффинности остальных молекул гематетрамерного Hb к O2.Таким образом, при связывании Hb с CO повышается аффинность Hb к O2, что препятствует отдаче кислорода в тканях и кривая связывания кислорода сдвигается влево. Из-за большой аффинности гемоглобина к угарному газу даже очень низкие парциальные давления CO приводят к связыванию значительного количества гемоглобина с CO с образованием HbCO. То, что у людей, которые находятся в условиях интенсивного уличного движения транспорта, или при курении, отравления угарным газом наступает редко, объясняется только медленным переходом угарного газа из альвеолярного газа в кровь. Двухвалентное железо гема (Fe2+) под действием сильных окислителей (нитраты, нитриты, анилинсодержащие вещества, бертолетовая соль и др.) может окисляться в трехвалентное (Fe3+). Гемоглобин с трехвалентным железом в геме называется метгемоглобин (MetHb, также хемиглобин). Метгемоглобин не может обратимо присоединять кислород и поэтому является для транспорта кислорода неактивным. Окисление железа гема ведет к увеличению аффинности остальных молекул гема к O2. Т.е., кривая связывания кислорода частично метгемоглобинизированным гемоглобином сдвинута влево. При помощи некоторых средств восстановления (дитионит) MetHb может опять восстановиться в гемоглобин, в геме которого железо опять будет двухвалентным. В организме восстановление метгемоглобина в гемоглобин происходит под влиянием фермента метгемоглобинредуктазы. Грудные дети, у которых метгемоглобинредуктаза еще недостаточно активна, особенно подвержены действию токсических веществ, окисляющих гемоглобин в метгемоглобин. ВОПРОСЫ ДЛЯ САМОПОДГОТОВКИ. 1.В каких формах кислород присутствует в крови? 2.Гемоглобин. Его роль в транспорте кислорода и углекислого газа. 3.Кривая диссоциация оксигемоглобина и ее изменения в зависимости от температуры, рН. 4.Транспорт углекислого газа от тканей к легким. 5. Транспорт ионов водорода от тканей к легким. 6. Виды гемоглобина. 4. РЕГУЛЯЦИЯ ДЫХАНИЯ Дыхательная система – система внешнего дыхания - это вегетосоматическая система с четкими морфологическими границами и конкретными механизмами деятельности, начиная от забора атмосферного воздуха, последующей транспортировки кислорода до клеток, окислительного обмена в тканях и кончая удалением углекислого газа из организма. Обеспечение клеток кислородом и удаление углекислого газа является главным, но не единственным назначением системы дыхания. Дыхательная система в различных звеньях связана с другими функциональными системами. Работа дыхательных мышц протекает в тесном взаимодействии и координации с общей моторной деятельностью организма. Процессы газообмена при дыхании тесно взаимосвязаны с функционированием сердечно-сосудистой системы. Вентиляторный аппарат обеспечивает работу обонятельного анализатора, а также звуковыражение у животных и речь у человека. Регулярное ритмическое поступление афферентации от рецепторов слизистой оболочки носа и рецепторных образований дыхательного аппарата, а также иррадиация ритмической активности из дыхательного центра оказывает огромное тоническое влияние на центральную нервную систему (ЦНС). Центральным звеном нервной регуляции дыхания является дыхательный центр (ДЦ). 4.1. Дыхательный центр. Впервые экспериментально достаточно точное местоположение дыхательного центра в продолговатом мозге было установлено в 1885 году Н.А. Миславским. Согласно Н.А. Миславскому, дыхательный центр расположен в ретикулярной формации дна четвертого желудочка по обеим сторонам срединного шва между верхушкой и основанием писчего пера, частично кнаружи, частично кнутри от ядер подъязычного нерва. В настоящее время идет интенсивное и всестороннее изучение этой структуры, определяющей жизненно важную функцию – дыхание. По современным представлениям, дыхательный центр рассматривается в первую очередь как совокупность нейронов, активность которых синхронна с фазами дыхательного цикла. Исходя из этого признака, предложены классификации нейронов. Наиболее популярной является классификация дыхательных нейронов по Bianchi et al. (1995). Согласно этой классификации, дыхательные нейроны в соответствии с паттерном их активности подразделяются на шесть основных групп: ранние инспираторные, инспираторные с нарастающим паттерном активности, поздние инспираторные, постинспираторные, экспираторные с нарастающим паттерном активности, преинспираторные. Дыхательные нейроны сосредоточены главным образом в пяти различных функционально-специфических отделах дыхательного центра: дорсальной дыхательной группе, вентральной дыхательной группе, пре-Бетцингеровом комплексе, комплексе Бетцингера и пневмотаксическом центре (рис. 9). Дорсальная дыхательная группа нейронов. Данная группа нейронов расположена в области ядра солитарного тракта. Большинство клеток составляют инспираторные нейроны различных типов: Rα , Rβ , Р-клетки и др. Rα-нейроны характеризуются дыхательной активностью, нарастающей по ходу вдоха, а также резкой ее остановкой в момент достижения объемного порога -выключения вдоха. Эти типы нейронов являются бульбо-спинальными, их аксоны образуют перекрест на уровне каудальной части продолговатого мозга, а затем направляются к спинальным мотонейронам инспираторных мышц. Паттерн активности Rβ-нейронов напоминает таковой у Rα-нейронов, но с некоторым отличием. Активность Rβ-нейронов не обрывается в конце вдоха, а сохраняется и в начале выдоха, постепенно убывая. 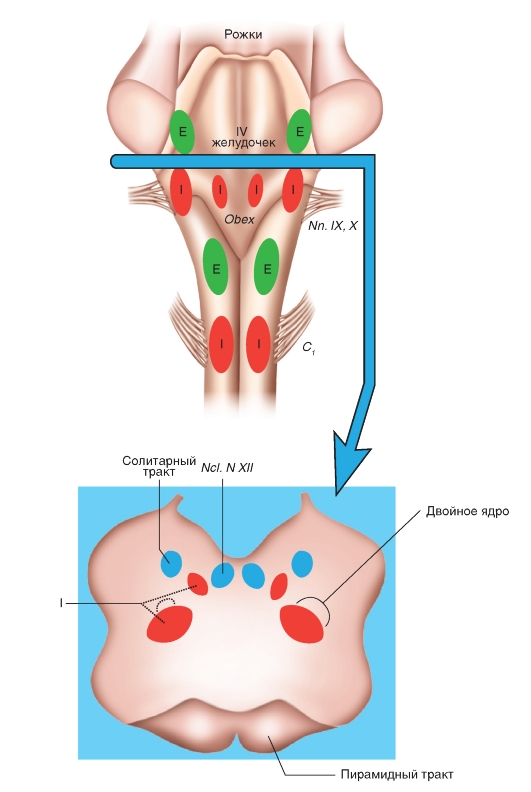 Рис.9. Дыхательный центр продолговатого мозга. Распределение респираторных нейронов в дыхательном центре продолговатого мозга. Вид на продолговатый мозг (medulla oblongata), инспираторные (I) и экспираторные нейронные группы (Е) и их приблизительное расположение. В действительности они находятся внутри ретикулярной формации и их трудно отграничить. Внизу: поперечный срез на данном уровне с инспираторными нейронными группами. Nn.IX,X – места выхода nn.Glossopharyngeus и Vagus. Ncl.N.XII – ядра подъязычного нерва. Рис. взят из кн. А.Г.Камкин, И.С.Киселева, 2012. Установлено, что в дополнение к общему источнику драйва, получаемому от генератора центральной инспираторной активности, к Rβ-нейронам приходит афферентация от рецепторов растяжения легких. Таким образом, Rβ-нейроны участвуют в реализации рефлекса Геринга-Брейера. Сравнительно недавно были описаны Р-клетки. Они получают информацию от рецепторов растяжения легких. Большинство нейронов дорсального ядра являются бульбоспинальными (у кошек от 50 до 80%). В отношении проприобульбарных нейронов дорсальной дыхательной группы известно, что многие из них проецируются в направлении вентролатеральных отделов дыхательного центра, а также к контрлатеральному ядру солитарного тракта. Предполагают, что эти проекции играют важную роль в передаче дыхательному центру интегрирующих афферентных импульсов, оказывающих влияние на дыхание. Ядро солитарного тракта отличается многообразием нейротрансмиттеров и нейропептидов, которые и обеспечивают его функции. Вентральная дыхательная группа. Нейроны вентральной дыхательной группы сосредоточены в двух симметричных колонках, вытянутых в рострально-каудальном направлении и расположенных в области n. ambigualis и n. retroambigualis. Вентральная дыхательная группа подразделяется на каудальный и ростральный отделы, различающиеся по функциональным характеристикам. Каудальный отдел вентральной дыхательной группырасположен вобласти n. retroambigualis. Этот отдел дыхательного центра состоит почти исключительно из бульбоспинальных экспираторных нейронов. У нейронов этой группы не обнаружено коллатералей на уровне продолговатого мозга. Предполагают, что они не имеют проекций к другим отделам бульбарного дыхательного центра. Данная группа экспираторных нейронов получает возбуждающий вход от более ростральных отделов дыхательного центра. Вероятно, активирующая афферентация поступает от экспираторных нейронов комплекса Бетцингера. Паттерн активности большинства экспираторных нейронов каудального отдела вентральной дыхательной группы характеризуется медленным нарастанием частоты импульсации на выдохе с максимумом в конце экспираторной фазы. Кроме экспираторных нейронов, в состав каудального отдела вентральной дыхательной группы входит небольшое количество инспираторных нейронов. Ростральный отдел вентральнойдыхательной группы расположен в области n. ambigualis и прилегающихпараамбигуальных структурах. Этот отдел состоит в основном из инспираторных нейронов. Здесь расположены бульбоспинальные премотонейроны инспираторных мышц, а также мотонейроны мышц гортани и некоторых мышц пищевода. В данном отделе также обнаружены проприобульбарные нейроны, которым приписывается координирующая роль в регуляции тонуса верхних дыхательных путей и инспираторных мышц. Паттерн активности инспираторных бульбоспинальных нейронов рострального отдела вентральной дыхательной группы характеризуется постепенным нарастанием частоты импульсации по ходу инспираторной фазы, аналогично паттерну активности диафрагмы и наружных межреберных мышц. В этом отделе дыхательного центра обнаружены нейроны с ранним инспираторным паттерном активности. Залпы таких нейронов характеризуются ранним началом, слегка опережающим активность диафрагмального нерва, быстрым достижением максимума с последующим снижением ее к концу инспираторной фазы. Считают, что ранние инспираторные нейроны являются источником ингибирования активности экспираторных премоторных клеток каудального отдела вентральной дыхательной группы и поздних инспираторных нейронов дорсальной дыхательной группы. В последнее время большое значение в генерации ритма дыхания придаетсяпре-Бетцингерову комплексу. У кошек он расположен в ростральной части n. ambigualis и вентролатеральной области ретикулярной формации каудальнее n. retrofacialis и ростральнее n. lateralis retikularis. При внеклеточной и внутриклеточной регистрации активности нейронов обнаружена область концентрации разнообразных типов дыхательных нейронов, в том числе двух типов преинспираторных нейронов, ранних инспираторных и постинспираторных нейронов. В данной структуре расположены те типы нейронов, которые согласно трехфазной теории ритмогенеза считаются необходимыми для генерации дыхательного ритма. Мнение о ведущей роли комплекса пре-Бетцингера в ритмогенезе опирается на некоторые экспериментальные данные. Так, в опытах in vitro удаление этого образования приводило к прекращению генерации дыхательного ритма в срезах ствола мозга. Локальные нарушения синаптической передачи в комплексе пре-Бетцингера также приводили к нарушению дыхательного ритма in vitro. Изучение нейронного состава комплекса пре-Бетцингера показало, что в данной структуре содержатся те типы нейронов, которые необходимы для генерации ритма в условиях in vitro: преинспираторные и пейсмекерные, а также нейроны, имеющие значения для переключения дыхательных фаз в этих условиях: преинспираторные и постинспираторные. Следует, однако, заметить, что вывод об исключительной роли данного образования в процессе генерации ритма дыхания является преждевременным и требует экспериментального исследования в условиях in vivo. Нейроны пре-Бетцингерова комплекса отличаются разнообразием медиаторов и их рецепторов. |