Физиология нервов, синапсов, мышц и рецепторов
 Скачать 23.71 Mb. Скачать 23.71 Mb.
|

Фа за де по ля ри за цииФа за де по ля ри за ции ПД обу слов ле на вхо дом в клет ку Na+. Дей ст ви тель но, де по ля ри за ция мо жет быть обу слов ле на толь ко вхо дом ка тио на,а из двух ка тио нов, уча ст вую щих в соз да нии био по тен циа лов (Na+ и K+), толь ко Na+ стре мит ся вой ти в клет ку. Из это го сле ду ет, что в фа зу де по ля ри за ции на трие вая про ни цае мость долж на рез ко по вы сит ся — то есть долж ны от крыть ся бы ст рые на трие вые ка на лы. Эти ка на лы, как уже го во ри лось, от кры ва ют ся в от вет на де по ля ри за цию. Сле до ва тель но, де по ля ри за ция яв ля ет ся един ст вен ным раз дра жи те лем, спо соб ным вы звать ПД. Итак, с од ной сто ро ны, де по ля ри за ция вы зы ва ет от кры ва ние на трие вых ка на лов, а с дру гой — вход Na+ по этим ка на лам вы зы ва ет де по ля ри за цию. Та ким об ра зом, вход Na+ в фа зу де по ля ри за ции ПД но сит са мо уси ли ваю щий ся ха рак тер: де по ля ри за ция при во дит к от кры ва нию на трие вых ка на лов; в ре зуль та те вхо дит Na+; вход Na+ при во дит к еще боль шей де по ля ри за ции; на рас таю щая де по ля ри за ция вы зы ва ет от кры ва ние но вых на трие вых ка на лов и т.д. Фа за ре по ля ри за цииНа пи ке ПД дви жу щие си лы, про ни цае мо сти и по то ки Na+ и K+ ме ня ют ся сле дую щим об ра зом.
Та ким об ра зом, фа за ре по ля ри за ции ПД обу слов ле на вы хо дом K+ на фо не пре кра щаю ще го ся вхо да Na+. Сле до вые по тен циа лыПо окон ча нии ПД час то воз ни ка ют так на зы вае мые сле до вые по тен циа лы, в боль шин ст ве слу ча ев обу слов лен ные тем, что во ро та ион ных ка на лов не ус пе ва ют вер нуть ся в со стоя ние, ха рак тер ное для по тен циа ла по коя. Ес ли сле до вой по тен ци ал по ло жи тель нее по тен циа ла по коя, то он на зы ва ет ся сле до вой де по ля ри за ци ей, ес ли от ри ца тель нее — сле до вой ги пер по ля ри за ци ей. Эти по тен циа лы в раз ных клет ках раз ные; в не ко то рых клет ках их нет во все, в дру гих мо жет быть не сколь ко сме няю щих друг дру га сле до вых по тен циа лов. В ка че ст ве при ме ра на рис. 1.10 при ве де на сле до вая ги пер по ля ри за ция. Вид но, что она обу слов ле на вре мен ным по вы ше ни ем ка лие вой про ни цае мо сти (из-за мед лен но го за кры ва ния по тен ци ал чув ст ви тель ных ка лие вых ка на лов) и, как след ст вие, сме ще ни ем мем бран но го по тен циа ла в сто ро ну EK. 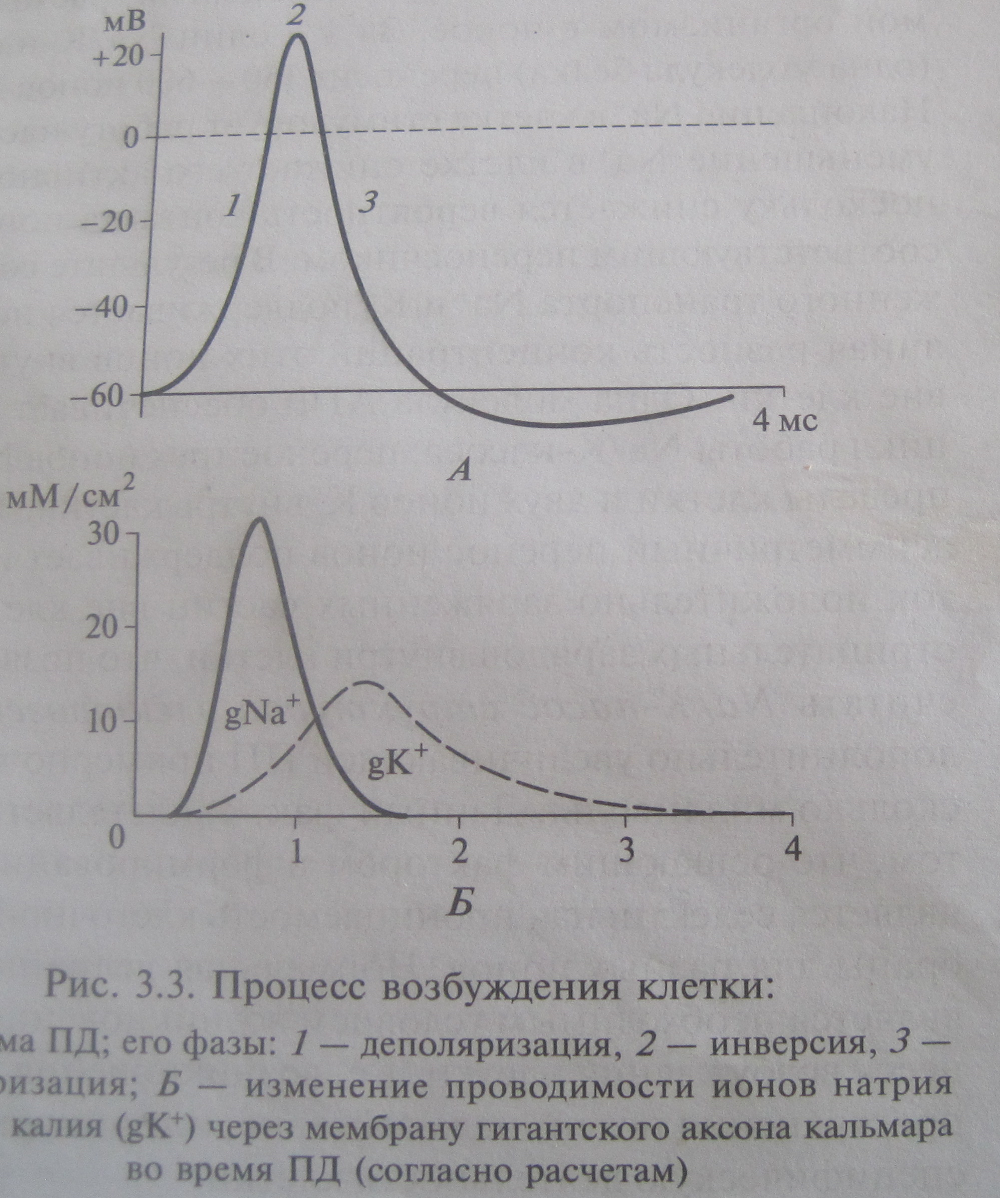
1. Во вре мя ло каль но го от ве та воз бу ди мость по вы ше на. При чи на: мем бран ный по тен ци ал при бли жа ет ся к Eкр. 2. Во вре мя фа зы де по ля ри за ции, пи ка ПД и пер вой тре ти фа зы ре по ля ри за ции воз бу ди мость па да ет до ну ля (ни ка кие раз дра жи те ли не спо соб ны вы звать по втор ный ПД). Это пе ри од аб со лют ной не воз бу ди мо сти, или аб со лют ной реф рак тер но сти — од но из клю че вых свойств ПД (см. вы ше, разд. «Фи зио ло ги че ский смысл био по тен циа лов»). При чи на: за кры ва ют ся инак ти ва ци он ные во ро та бы ст рых на трие вых ка на лов, и в этих ус ло ви ях ни ка кие раз дра жи те ли не спо соб ны вы звать от кры ва ние этих ка на лов. 3. На чи ная от вто рой тре ти фа зы ре по ля ри за ции воз бу ди мость по сте пен но вос ста нав ли ва ет ся, хо тя и ос та ет ся сни жен ной по срав не нию с ис ход ной (ПД мо жет быть вы зван, но толь ко силь ны ми раз дра жи те ля ми). Это пе ри од от но си тель ной не воз бу ди мо сти, или от но си тель ной реф рак тер но сти. При чи на: по сте пен но вновь от кры ва ют ся инак ти ва ци он ные во ро та бы ст рых на трие вых ка на лов. Ес ли сле до вых по тен циа лов нет, то воз бу ди мость вос ста нав ли ва ет ся поч ти од но вре мен но с окон ча ни ем фа зы ре по ля ри за ции (рис 1.14, А). Ес ли же име ют ся сле до вые по тен циа лы, то из ме не ния воз бу ди мо сти бо лее слож ны. В ча ст но сти, во вре мя сле до вой ги пер по ля ри за ции воз бу ди мость по ни же на, по это му пе ри од от но си тель ной реф рак тер но сти длит ся доль ше — до окон ча ния сле до вой ги пер по ля ри за ции (рис. 1.14, Б). При чи ны: 1) мем бран ный по тен ци ал сме щен в сто ро ну бо лее от ри ца тель ных зна че ний; 2) ка лие вая про ни цае мость по вы ше на, и по это му Eкр сме щен в сто ро ну бо лее по ло жи тель ных зна че ний (рис. 1.13, В). Обе эти при чи ны при во дят к рос ту по ро га де по ля ри за ции, и, сле до ва тель но, сни же нию воз бу ди мо сти. Ка ко вы бы ни бы ли осо бен но сти из ме не ний воз бу ди мо сти в от дель ных ней ро нах, глав ной и важ ней шей осо бен но стью этих из ме не ний яв ля ет ся на ли чие аб со лют ной реф рак тер но сти: поч ти все вре мя, по ка длит ся ПД, по втор ный ПД вы звать не воз мож но. От дли тель но сти пе рио да аб со лют ной реф рак тер но сти за ви сит, сколь ко ПД в 1 се кун ду мо жет воз ник нуть в той или иной клет ке. На при мер, ес ли реф рак тер ность со став ля ет 1 мс, то мак си маль ная час то та ПД рав на око ло 1000 в 1 с, то есть 1000 Гц. Эта мак си маль ная час то та им пуль сов, ко то рую мо жет вос про из ве сти та или иная ткань, клет ка или часть клет ки, яв ля ет ся ме рой ла биль но сти («функ цио наль ной под виж но сти тка ни»). 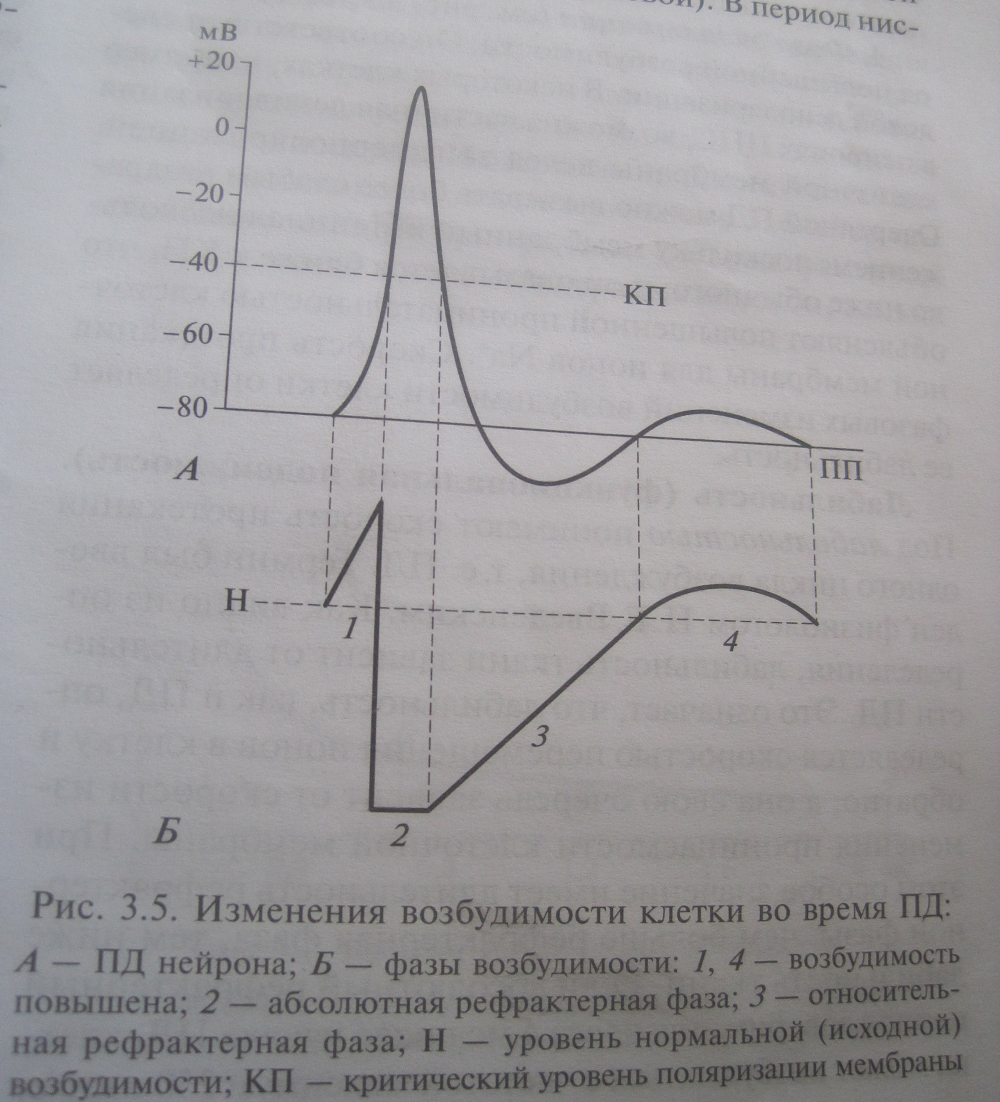 
Возбудимость клетки оценивается несколькими параметрами.
Ак ко мо да ция — это сни же ние воз бу ди мо сти при умень ше нии кру тиз ны (ско ро сти на рас та ния) раз дра жи те ля. При чи на ак ко мо да ции — та же, что и дли тель ной де по ля ри за ции: при мед лен но на рас таю щей де по ля ри за ции ус пе ва ют раз вить ся на трие вая инак ти ва ция и ка лие вая ак ти ва ция. Лабильность – функциональная подвижность, скорость протекания отдельных циклов возбуждения в возбудимых тканях. Мера лабильности – максимальное количество импульсов, которое может воспроизвести ткань в единицу времени в соответствии с ритмом возбуждения. Лабильность определяется скоростью перемещения ионов в клетку и обратно, и она в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. Чем больше рефрактерная фаза, тем ниже лабильность ткани.
Нервные волокна – отростки нейронов, с помощью которых осуществляется связь нейронов между собой, а также с иннервируемыми клетками и рецепторами. Оболочку безмиелиновых волокон образуют швановские клетки, в которые погружаются осевые цилиндры нейронов. Оболочку миелиновых волокон образуют в периферической НС леммоциты, а в ЦНС олигодендроциты. Миелиновая оболочка влияет на электрические свойства волокна, способствует увеличению скорости проведения возбуждения, выполняет трофическую функцию (регуляция метаболизма, рост осевого цилиндра) Плазматическая мембрана определяет пассивные электрические свойства волокна (сопротивление и емкость мембраны). Распределение и концентрация электроуправляемых каналов влияет на возбудимость и проводимость нервного волокна. Нейрофибрилы и микротрубочки обеспечивают аксональный транспорт. Появление разности потенциалов между возбужденными и невозбужденными участками мембраны. =>возникновение локальных токов между возбужденными и невозбужденными участками мембраны=>генерация новых ПД в соседних участках волокна. Законы проведения возбуждения по нервному волокну
Если в эксперименте нанести раздражение в любой участок нервного волокна, то возбуждение регистрируется как в проксимальном, так и в дистальном участке нерва
Синапс – специализированная структура, обеспечивающая передачу сигнала от клетки к клетке с помощью химических веществ или электрического поля. В состав синапса входит пре и постсинаптическая мембраны и синаптическая щель. В области пресинаптической мембраны находятся многочисленные везикулы, которые содержат молекулы медиатора ацетилхолина. Везикулы концентрируются в особых областях- активных зонах. Постсинаптическая мембрана образует постсинаптические складки, на гребнях которых располагаются АХ – рецепторы. Синаптическая щель заполнена межклеточной жидкостью, в ней располагается базальная пластинка, которая содержит различные белки. Они способствуют плотному сцеплению пре и постсинаптических мембран. Базальная мембрана содержит фермент ацетилхолинэстераза, котрая разрушает высвобожденный ацетилхолин и готовит синапс к проведению очередного сигнала. Механизм синаптической передачи.
Особенности проведения в синапсе по сравнению с нервным волокном.
7. Скелетная мышца: значение отдельных структурных элементов мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы и двигательного аппарата организма, классификация двигательных единиц, физиологические свойства скелетной мышцы и ее функции Структурной и функциональной единицей мышцы, является мышечное волокно, представляющее собой сильно вытянутое многоядерное волокно. Мышечное волокно имеет оболочку сарколемму. В саркоплазме мышечного волокна имеются многочисленные ядра, митохондрии, растворимые белки, капельки жира, гранулы гликогена, фосфатсодержащие вещества, другие малые молекулы, а также ионы. Там же расположены многочисленные мембраны. Они включены в состав поперечных Т – трубочек, пересекающих мышечное волокнои связывающих сарколемму с внутриклеточным пространством и саркоплазматическим ретикулумом. Последний представляет собой систему связанных друг с другом систем и канальцев. Т – система и ретикулум обеспечивают функциональное согласование процессов возбуждения клеточной мембраны со специфической активностью сократительного аппарата миофибриллы. Через Т – трубочки может происходить выделение продуктов обмена в межклеточное пространство и далее в кровь. Внутри мышечного волокна тянется масса миофибрилл, с которыми связана способность мышц к сокращению. Двигательная единица – комплекс мотонейрона со всеми его коллатералями и группой иннервируемых им мышечных волокон. Двигательные единицы подразделяют на быстрые и медленные. Медленные мотонейроны низкопороговые, малоутомляемые. Быстрые мотонейроны высокопороговые, утомляемые, не способны к длительному поддержания высокочастотного разряда. Физиологические свойства скелетной мышцы. 1.Возбудимость. Способность мышцы отвечать на действие раздражителя самой мышцы или двигательного нерва изменением физиологических свойств и возникновением возбуждения. 2.Проводимость. Способностъ проводить возбуждение, возникшее в каком-либо участке мышечного волокна, по всему волокну. 3.Рефрактерность. Временное снижение возбудимости мышцы, которое возникает в результате возбуждения. 4.Лабильность. Количество возбуждений за единицу времени, зависящее от уровня обменных процессов. 5.Сократимость. Способность изменять свою длину или напряжение при возбуждении. Это основная функция скелетном мышцы. В период относительного покоя скелетные мышцы полностью не расслаблены, а умеренно напряжены. Такое состояние называется мышечным тонусом и объясняется редкими импульсами от двигательных нейронов, которые попеременно возбуждают нейромоторные единицы. При изотоническом сокращении укорачивается мышечное волокно, а напряжение не изменяется; при изометрическом сокращении длина мышцы не изменяется, а напряжение возрастает. При этом мышцы выполняют следующие функции: 1) обеспечивают определенную позу тела человека; 2) перемещают тело в пространстве; 3) перемещают отдельные части тела относительно друг друга; 4) являются источником тепла, выполняя терморегуляционную функцию. 8. Механизм сокращения и расслабления скелетной мышцы: значение потенциала действия ионов кальция, тропонина и тропомиозина, миозиновых мостиков, АТФ. |