Физиология нервов, синапсов, мышц и рецепторов
 Скачать 23.71 Mb. Скачать 23.71 Mb.
|

|
Процессы торможения в ЦНС: экспериментальные данные (И.М.Сеченов, Гольц, Мегун), механизм постсинаптического и пресинаптического торможений, их разновидности (показать с помощью схемы), значение торможения. Торможение – активный нервные процесс , результатом которого является ослабление или прекращение возбуждения. Торможение в ЦНС открыл Сеченов. В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Латентное время значительно увеличивается если на зрительный бугор предварительно положить кристаллик поваренной соли. Гольц обнаружил проявления торможения у спинальной лягушки. Механическое раздражение кончиков пальцев одной лапки существенно удлиняет латентное время сгибательного рефлекса другой при погружении ее в раствор кислоты. Мегун доказал наличие специальных тормозных структур в продолговатом мозге. В опытах на кошках при изучении разгибательного рефлекса он обнаружил, что раздражение медиальной части РФ продолговатого мозга тормозит рефлекторную деятельность спинного мозга. Особенности и механизм постсинаптического торможения. Постсинаптическое торможение обнаружил и описал Дж. Экклс при регистрауции потенциалов мотонейронов спинного мозга у кошки во время сокращения и расслабления мышцы в ходе реализации соответствующих рефлекторных актов. При рефлекторном расслаблении мышц на мотонейронах регистрируется гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона и угнетающий его способность реагировать на возбуждающие влияния. Это ТПСП. Амплитуда ТПСП 1 – 5 мВ. Он способен суммироваться. Тормозные синапсы постсинаптического торможения локализуются в основном на теле нейрона. Механизм постсинаптического торможения. Возбудимость клетки во время ТПСП понижается, поскольку увеличивается пороговый потенциал, т.к. КП остается на прежнем уровне, а мембранный потенциал возрастает. ТПСП возникает под действием аминокислоты глицина и ГАМК. В спинном мозге глицин выделяется в синапсах особых тормозных клеток (клетки Реншоу), образуемых этими клетками на мембране постсинаптического нейрона. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для хлора, при этом хлор поступает в клетку согласно концентрационному градиенту, в результате развивается гиперполяризация. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа хлора и выхода калия из клетки. Активация ГАМК1 рецепторов ведет к непосредственному увеличению проницаемости мембраны для хлора. Активация ГАМК2 рецепторов реализуется с помощью вторых посредников (цАМФ), при этом повышается проницаемость для калия. ТПСП могут возникать вследствие уменьшения проницаемости мембраны для натрия, что сопровождается гиперполяризацией клеточной мембраны. Разновидности постсинаптического торможения. Выделяют возвратное, латеральное, параллельное и прямое (реципрокное) постсинаптическое торможение. Возвратное постсинаптическое торможение – такое торможение, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. Типичным примером является торможение в мотонейронах спинного мозга. Параллельное торможение возбуждение блокирует само себя, за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением. Латеральное тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями, в результате в соседних клетках развивается глубокое торможение. Прямое торможение вызывает угнетение центра антагониста. Возбуждающие импульсы поступают в клетку мышцы сгибателя, а через тормозную клетку к центру мышцы антагониста. Пресинаптическое торможение. Было выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных альфа мотонейронов. Однако опережающее раздражение афферентов сухожильных рецепторов мышц антагонистов предотвращало возбуждение активируемых альфа мотонейронов. Не изменялись мембранный потенциал и возбудимость исследуемых мотонейронов, либо регистрировался низкоамплитудный ВПСП, не способный вызвать ПД. Механизм пресинаптического торможения. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях медиатором является ГАМК, который действует на ГАМК1 рецепторы. Под действием ГАМК тормозных нейронов и последующего повышения проницаемости мембраны для хлора ионы хлора начинают выходить согласно электрическому градиенту. Это приводит к деполяризации пресинаптических терминалей и ухудшению их способности проводить импульсы. Разновидности пресинаптического торможения. Можно разделить на две группы: в первой блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение); во второй блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое). Роль различных видов торможения. Торможение выполняет охранительную ролью. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС. Большое значение имеет для обработки поступающей в ЦНС информации. Торможение параллельных путей способствует выдеоению существенных сигналов из фона. Является важным фактором координационной деятельности ЦНС. 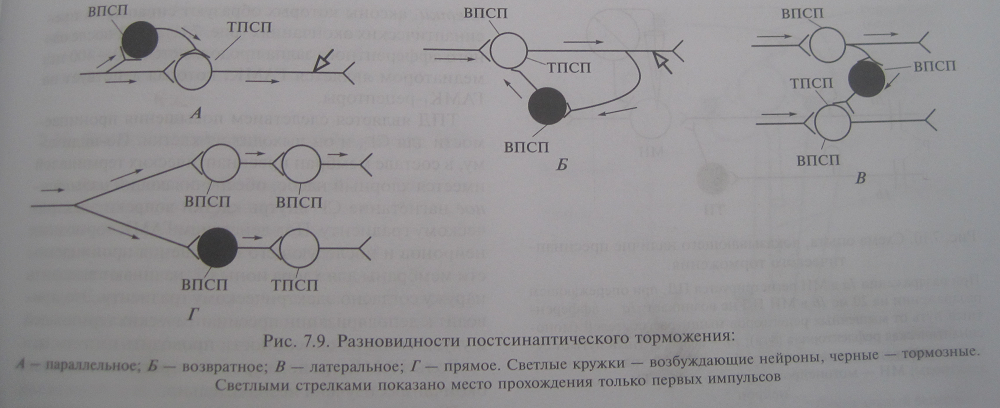 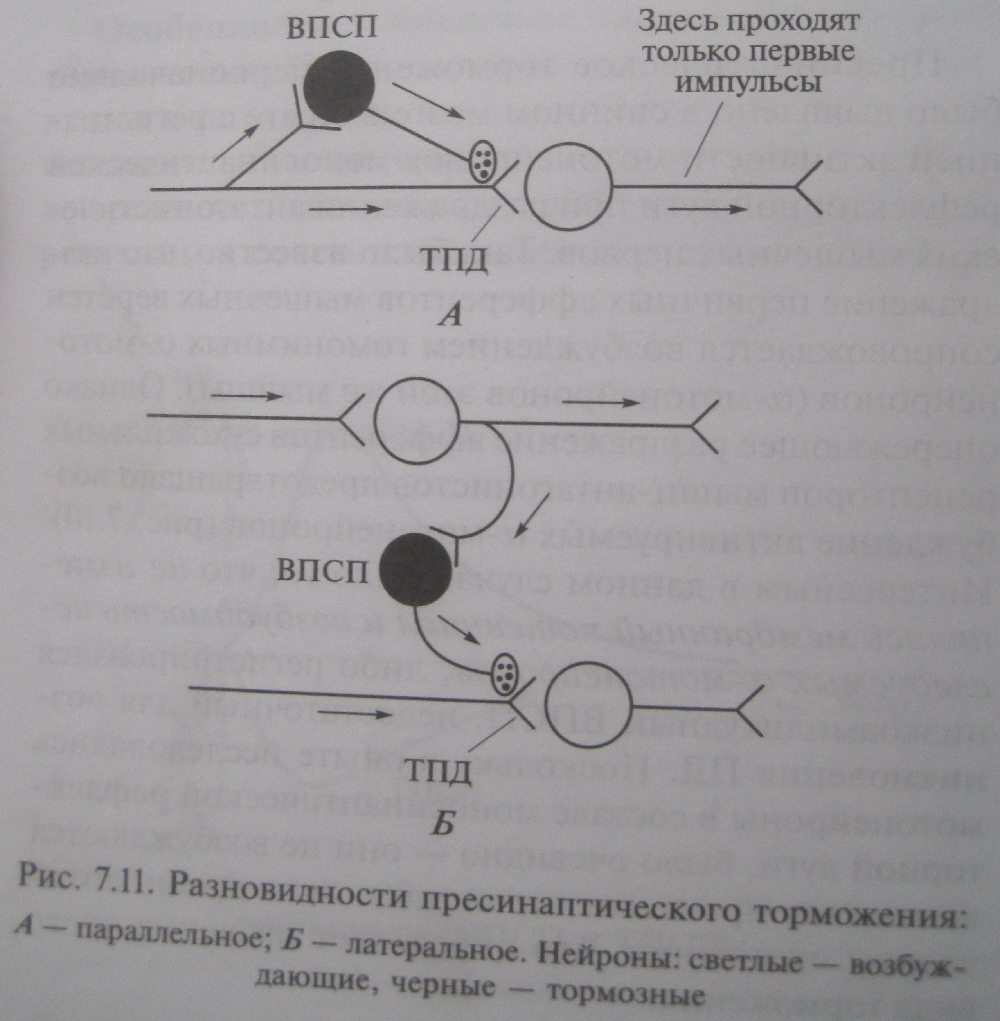
Координационная деятельность ЦНС. Это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Основой является взаимодействие процессов возбуждения и торможения. Координационная деятельность обеспечивается несколькими факторами.
Прямая связь – управление другим центром или рабочим органом с помощью посылки к ним эфферентных импульсов. Обратная связь (обратная афферентация) – управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них самих. Возвратная связь обеспечивает торможение нейронов вслед за их возбуждением. Реципрокная (сочетанная) связь обеспечивает торможение центра антагониста при возбуждении центра агониста. Принцип модульной (ансамблевой) структурно – функциональной организации ЦНС: каждый модуль представляет собой совокупность повторяющихся локальных нейронных связей, обрабатывающих и передающих инфу с помощью внутренних и внешних связей. Основным функциональным признаком модульной организации является локальный синергизм реакций нейронов центральной части ансамбля, окруженной зоной заторможенных нейронов – тормозная оконтовка.
Соматические нейроны спинного мозга. Нейроны спинного мозга классифицируют по принадлежности к отделам НС – соматические и вегетативные; по назначению – эфферентные, афферентные, вставочные, ассоциативные; 3)по влиянию – возбуждающие и тормозные. Эфферентные нейроны относятся к соматической нс, являются эффекторными, поскольку иннервируют непосредственно рабочие органы – эффекторы (скелетные мышцы), их называют мотонейроны. Различают альфа и гама мотонейроны. Альфа мотонейроны иннервируют экстрафузальные мышечные волокна (скелетная мускулатура), их аксоны характеризуются высокой скоростью проведения возбуждения. Они подразделяются на две подгруппы альфа1 мотонейроны – быстрые, иннервирующие белые мышечные волокна, и альфа2 – медленные, иннервирующие красные мышечные волокна. Гамма – мотонейроны рассеяны среди альфа мотонейронов, их активность регулируется нейронами вышележащих отделов ЦНС. Они обладают спонтанной активностью, иннервируют интрафузальные мышечные волокна мышечного веретена (мышечного рецептора). Афферентные нейроны локализуются в спинальных ганглиях и ганглиях черепных нервов. Их отростки, проводящие афферентную импульсацию от мышечных, сухожильных и кожных рецепторов, вступают в ствол мозга или соответствующие сегменты спинного мозга и образуют синаптические контакты либо на альфа мотонейронах, либо на вставочных нейронах. Вставочные нейроны устанавливают связь с мотонейронами спинного мозга чувствительных нейронов. Они обеспечивают связь спинного мозга с ядрами ствола , а через них с корой большого мозга. Ассоциативные нейроны образуют собственный аппарат спинного мозга, устанавливающий связь между сегментами и внутри одного сегмента. Участвует в поддержании позы, тонуса мышц, движений. Нейроны РФ. РФ спинного мозга состоит из тонких перекладин серого вещества, пересекающихся в различных направлениях, ее нейроны имею большое количество отростков. РФ обнаруживается на уровне шейных сегментов между передними и задними рогами, а на уровне верхнегрудных сегментов – между боковыми и задними рогами в белом веществе, примыкающем к серому. Спинальные нейроны вегетативной нервной системы. Неуроны симпатической нервной системы являются вставочными. Расположены в боковых рогах грудного, поясничного и частично шейного отделов. Фоноактивны. Нейроны парасимпатической НС также вставочные и фоноактивные, локализуются в сакральном отделе спинного мозга (S2 – S4). Центры спинного мозга. Различные центры симпатического отдела НС локалихзованы в: центр зрачкового рефлекса – C8 – Th12; центр регуляции деятельности сердца – Th1 – Th5; слюноотделения – Th2 – Th4; регуляции функции почек – Th5 – L3; сегментарно расположены центры регулирующие функции половых желез и сосудов, гладких мышц внутренних органов, центры пиломоторных рефлексов. Парасимпатическую иннервацию (S2 – S4) получают все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы. Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу скелетную мускулатуру шеи, диафрагмы, верхних конечностей, туловища и нижних конечностей. Проводниковая функция спинного мозга осуществляется нисходящих и восходящих путей. Афферентная инфа поступает в спинной мозг в основном через задние корешки, эфферентная импульсация в передних корешках и регуляция функций различных органов и тканей организма осуществляется через передние корешки (закон Белла – Мажанди). Все афферентные входы в спинной мозг несут инфу от трех групп рецепторов: 1) кожных – болевых, температурных, прикосновения, давления, щекотки, вибрации; 2) проприорецепторов мышц (мышечных веретен), сухожилий (рецепторов Гольджи), надкостницы и оболочек суставов; 3) от рецепторов внутренних органов – висцерорецепторов (механо и хеморецепторов). Медиатором первичных афферентных нейронов является глутамат, модулятором – субстанция П, энкефалин, ВИП. Значение афферентной импульсации заключается в следующем: 1) участвует в координационной деятельности ЦНС по управлению скелетной мускулатурой; при выключении афферентной импульсации от рабочего орган управление им становится несовершенным; 2) участвует в процессах регуляции функций внутренних органов; 3) поддерживает тонус ЦНС, при выключении афферентной импульсации происходит уменьшение суммарной тонической активности ЦНС; 4) афферентная импульсация несет в вышележащине отделы ЦНС инфу об изменениях окружающей среды. Тонус мышц у спинального организма. Тонус формируют статические тонические рефлексы мышц сгибателей и разгибателей. Сразу после перерезки или при повреждении спинного мозга у человека наблюдается мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной является выключений влияния на спинной мозг вышележащих отделов ЦНС. Фазные сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются. Гипертонус имеет рефлекторную природу, он развивается вследствие афферентной импульсации от мышечных рецепторов. Имеется два фактора, обеспечивающих афферентную импульсацию от мышечных рецепторов по задним корешкам спинного мозга, которая ведет к возбуждению альфа мотонейронов спинного мозга и сокращению скелетных мышц. 1) спонтанная активность мышечных рецепторов, поступление афферентных импульсов к альфа мотонейронам, последние возбуждаются и посылают импульсы к мышцам, повышая тонус. 2) спонтанная активность гамма мотонейронов. Возбуждение гамма мотонейронов вызывает возбуждение и сокращение иннервируемых ими интарфузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его прикреплены к скелетной мышце. Вследствие натяжения мышечного рецептора раздражаются и возбуждаются афферентные окончания (проприорецепторы), импульсы от которых поступают к альфа мотонейронам и возбуждают их. Альфа мотонейроны посылают импульсы к скелетной мышце, вызывая ее постоянное (тоническое) сокращение.
Классификация соматических рефлексов спинного мозга. По иду рецепторов, раздражение которых вызывает рефлекс: проприорецептивные, висцерорецептивные, кожные. Последниф являются защитными. Рефлексы, возникающие с проприроепторов, участвуют в акте ходьбы и регуляции мышечного тонуса, что важно для поддержания позы организма. Висцеререцептивеные рефлексы возникают с интерорецепторов и проявляются в сокращении мышц брюшной стенки, грудной клетки и разгибателей спины. По органам (эффекторам рефлекса): рефлексы конечностей, брюшные, области таза. Рефлексы конечностей могут быть фазными и тоническими. Фазные рефлексы подразделяются на сгибательные, разгибательные и ритмические.
Тонические рефлексы конечностей включают две группы: сегментарные и надсегментарные.
Брюшные рефлексы. Верхний, нижний, средний брюшные рефлекс являются фазными. Они выражаются в сокращениях соответствующих участков мускулатуры стенки живота. Это защитные рефлексы. Рефлексы области таза. К ним относятся кремастерный (яичковый) и анальный рефлексы. Оба фазные. Шагательный рефлекс. Рефлекс вызывают однократным раздражением кожи конечности. Он выражается в сгибании этой конечности с одновременным разгибанием противоположной задней конечности – перекрестный разгибательный рефлекс. Затем согнутая конечность разгибается, опускается вниз, а разогнутая сгибается и поднимается вверх и т.д. Значение проприорецепторов в шагательном рефлексе. Мышечные веретена (мышечные проприорецепторы) расположены параллельно скелетной мышце: своими концами крепятся к соединительно тканной оболочке пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия волокон соединительной ткани. Поэтому, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению. Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой. Вокруг средней части мышечного веретена обвивается несколько раз окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим афферентным волокном, которое также обвивает в виде спирали мышечный рецептор, но периферические его участки. Импульсы от мышечных рецепторов возбуждают нейроны своего двигательного центра и тормозят нейроны центра антагониста. Ближе к концам мышечных веретен подходят двигательные нервные окончания, аксоны гамма мотонейронов спинного мозга. Их импульсация вызывает сокращение интарфузальных мышечных волокон, что ведет к возбуждению рецептора. Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно – мышечного соединения. Они представляют собой безмиелиновыее окончания толстого миелинового афферентного волокна. Крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение и возбуждение при натяжении сухожилий. Сухожильные рецепторы посылают в мозг инфу о том, что мышца сокращена, тогда как мышечные рецепторы сигнализируют о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра антагониста. Механизм шагательного рефлекса. Поочередное сокращение и расслабление скелетных мышц запрограммированы в спинном мозге, важную роль играет импульсация от проприорецепторов в двигательный центр к каждой мышце. Когда мышца расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим альфа мотонейронам спинного мозга и возбуждают их. Далее альфа мотонейроны посылают импульсы к той же мышце, что ведет к ее сокращению. Как только мышца сократилась, возбуждение мышечных веретен прекращается или сильно уменьшается и начинают возбуждаться сухожильные рецепторы. Импульсы от последних поступают в свой спинальный центр, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение альфа мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление снова возбуждает мышечные рецепторы и альфа мотонейроны и мышца вновь сокращается. Шагательные координированные движения возможны у спинального животного в отсутствие обратной афферентации проприорецепторов, благодаря наличию генератора локомоторного цикла (каждая конечность может совершать самостоятельные движения) при этом важную роль играют межсегментарные связи на уровне спинного мозга. 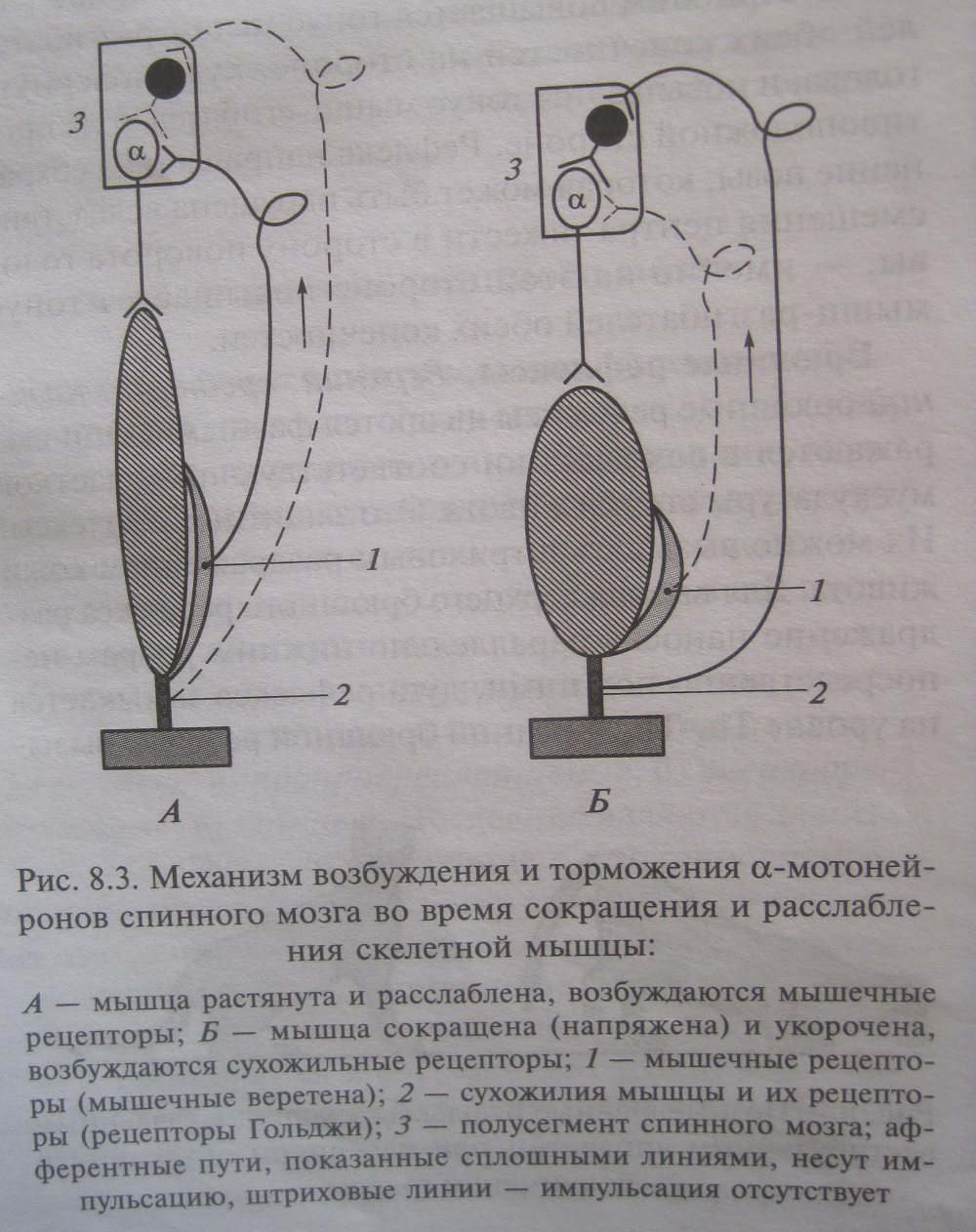 |