Физиология нервов, синапсов, мышц и рецепторов
 Скачать 23.71 Mb. Скачать 23.71 Mb.
|

|
Продолговатый мозг и мост: центры и соответствующие им рефлексы, их отличия от рефлексов спинного мозга, состояние мышечного тонуса бульбарного животного, схема, объясняющая механизм его происхождения. Непосредственным продолжением вверх спинного мозга у всех позвоночных животных и человека является продолговатый мозг. Вместе с варолиевым мостом продолговатый мозг является производным ромбовидного мозга (rhombencephalon). Одновременно он является частью ствола мозга (truncus encephalicus). Ствол - это осевые структуры головного мозга, включающие продолговатый мозг, мост, средний и промежуточный мозг. В продолговатом мозге и мосту по сравнению со спинным мозгом нет четкой сегментации серого и белого вещества. Скопления нейронов здесь формируют ядра, обеспечивающие сложную рефлекторную деятельность. В продолговатом мозге и варолиевом мосту (в дорзальной их части) находится большая группа ядер черепных нервов (с V по XII пары), которые образуют массу серого вещества ромбовидной ямки. Эти ядра являются жизненно важными функциональными центрами. На дне IV желудочка в продолговатом мозге находится дыхательный центр, состоящий из центра вдоха и центра выдоха. Нейроны этих центров посылают импульсы к дыхательным мышцам через мотонейроны спинного мозга, контролируя правильное чередование фаз дыхательного цикла. Рядом лежит сосудодвигательный центр (контролирует тонус стенок сосудов и уровень АД) и сердечно-сосудистый центр (согласует состояние сосудов с работой сердца). В продолговатом мозге также лежат пищеварительные центры: 1 - моторные (жевания, глотания, моторики желудка и части кишечника), 2 - секреторные (слюноотделения, желудочной секреции, выделения соков тонкой кишки, поджелудочной железы и др.). Здесь же находятся центры защитных рефлексов (чихания, кашля, рвоты, мигания, слезоотделения). Таким образом, биологическая роль продолговатого мозга заключается в регуляции постоянства состава внутренней среды организма (гомеостаза). Он также осуществляет более тонкие приспособительные реакции организма к внешней среде, чем спинной мозг. Кроме ядер черепных нервов в продолговатом мозге имеются переключательные чувствительные ядра. В толще олив лежат крупные нижние оливные ядра, а также медиальные и верхние добавочные оливные ядра. На задней поверхности в одноименных бугорках залегают тонкое ядро Голля и клиновидное ядро Бурдаха. Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса мышц. Импульсы от вестибулярных ядер (Дейтерса и Бехтерева) усиливают тонус мышц разгибателей, что необходимо для организации позы. Неспецифические отделы продолговатого мозга (ядра РФ) наоборот снижают тонус мышц, в том числе и разгибателей. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела (установочных рефлексов). Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется за счет: 1 - простых сегментарных рефлексов (защитный мигательный, слезоотделения, движений ушной раковины, кашля, чихания, рвоты), 2 - лабиринтных рефлексов (распределение тонуса между отдельными группами мышц и установки определенной позы), 3 - установочных рефлексов (поддержания позы и рабочих движений), 4 - вегетативных рефлексов (дыхания, кровообращения, пищеварения). Проводниковая функция осуществляется путем проведения: 1 - восходящих волокон от спинного мозга к коре полушарий, 2 - нисходящих волокон от коры полушарий к спинному мозгу, 3 - собственных проводящих пучков продолговатого мозга и моста, соединяющих ядро и оливу вестибулярного нерва с мотонейронами спинного мозга. Продолговатый мозг принимает чувствительные волокна от рецепторов мимических и жевательных мышц, мышц шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, полости носа и рта, от рецепторов органов слуха и равновесия, от рецепторов гортани, трахеи, легких, интерорецепторов ЖКТ и сердечно-сосудистой системы. Здесь волокна переключаются на другие нейроны, образуя путь в таламус и кору полушарий. Восходящие пути кожно-мышечной чувствительности перекрещиваются на уровне продолговатого мозга так же, как и большая часть пирамидных (двигательных) путей. Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего мозга. Бульбарный организм характеризуется децеребрационной ригидностью. В таком организме сохранены шейные и лабиринтные тонические рефлексы, но отсутствуют выпрямительные рефлексы и произвольные движения. 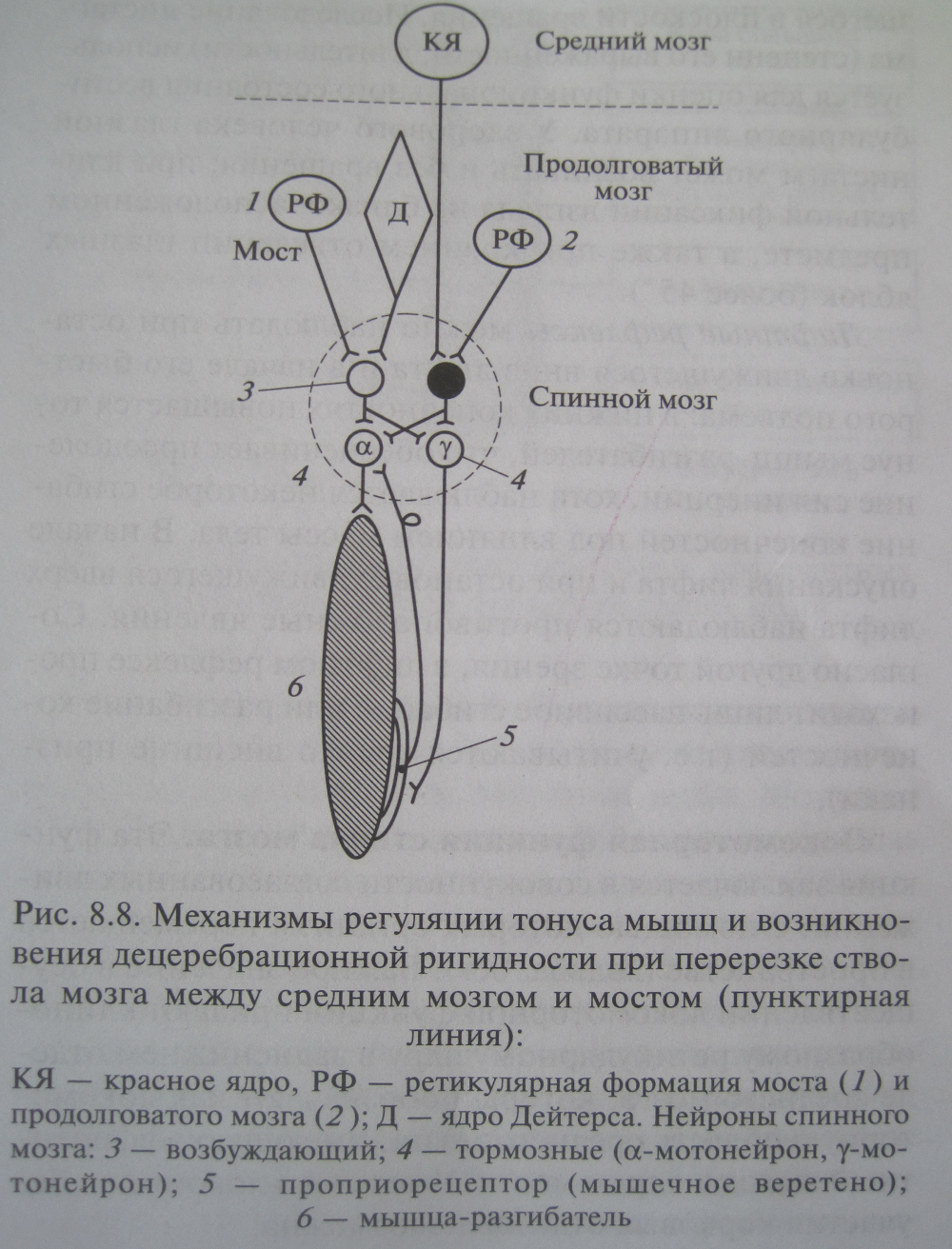
Средний мозг — отдел головного мозга, древний зрительный центр. Включен в ствол головного мозга. Вентральную часть составляют массивные ножки мозга, основную часть которых занимают пирамидные пути. Между ножками находится межножковая ямка, из которой выходит III (глазодвигательный) нерв. В глубине межножковой ямки — заднее продырявленное вещество. Дорсальная часть — пластинка четверохолмия, две пары холмиков, верхние и нижние. Верхние, или зрительные холмики несколько крупнее нижних (слуховых). Холмики связаны со структурами промежуточного мозга — коленчатыми телами, верхние — с латеральными, нижние — с медиальными. Внутри нижних холмиков находятся слуховые ядра, туда идет латеральная петля. Вокруг сильвиева водопровода — центральное серое вещество. В глубине покрышки среднего мозга (под четверохолмием) находятся ядра глазодвигательных нервов, красные ядра, чёрное вещество, ретикулярная формация. В латеральных отделах среднего мозга в него входят верхние мозжечковые ножки, которые, постепенно погружаясь в него, образуют перекрест у средней линии. Дорсальная часть среднего мозга, расположенная кзади от водопровода, представлена крышей с ядрами нижних и верхних холмиков. Ядра нижних холмиков играют существенную роль в реализации функции слуха и формировании сложных рефлексов в ответ на звуковые раздражения. Ядра участвуют в осуществлении «автоматических» реакций, связанных со зрительной функцией, то есть безусловных рефлексов в ответ на зрительные раздражения. Кроме того, эти ядра координируют движения туловища, мимическую реакцию, движения глаз, головы, ушей и проч. в ответ на зрительные стимулы. Осуществляются эти рефлекторные реакции благодаря покрышечно-спиномозговому и покрышечно-бульбарному путям. Вентральнее от верхних и нижних холмиков крыши находится водопровод среднего мозга, окружённый центральным серым веществом. В нижнем отделе покрышки среднего мозга располагается ядро блокового нерва, а на уровне среднего и верхнего отделов — комплекс ядер глазодвигательного нерва. Важными образованиями среднего мозга являются также красные ядра и чёрная субстанция. В красных ядрах оканчиваются волокна передних мозжечковых ножек, корково-красноядерные волокна и волокна из образований стриопаллидарной системы. В красном ядре начинаются волокна красноядерно-спинномозгового, а также красноядерно-оливного путей, волокна, идущие в кору большого мозга. Таким образом, красное ядро является одним из центров, участвующих в регуляции тонуса и координации движений. В чёрном веществе берет своё начало нигростриарный путь, дофаминэргические волокна которого управляют функцией стрии. Основание ножки среднего мозга состоит из волокон, которые соединяют кору большого мозга и другие образования конечного мозга с нижележащими образованиями мозгового ствола и спинного мозга. Большая часть основания занята волокнами пирамидного пути. Средний мозг выполняет следующие функции: центр ориентировочного рефлекса центр позы центр обработки первичной информации (зрение, слух) регулирующую в продолжительности актов жевания и глотания Черная субстанция ограничивает влияние норадренергической системы, активирует продукцию оксида азота в головном мозге, увеличивает устойчивость к стрессорным повреждениям. Нейроны черного вещества оказывают модулирующее влияние на фоновую активность нейронов полосатого тела. Также оказывает тормозящее действие на нейроны таламуса. Мезенцефальный организм (нарушение связи между средним и промежуточным мозгом) имеет нормальный мышечный тонус, сохраняются все тонические рефлексы, но произвольные движения отсутствуют. Тонические рефлексы организма Магнус разделил на две группы: статические (перераспределение мышечного тонуса при изменении положения тела в пространстве, или отдельных его частей относительно друг друга, например восстановление нарушенной позы) и статокинетические (перераспределение мышечного тонуса при перемещении организма в пространстве, например при беге, подъеме в лифте). Статические рефлексы – тонические рефлексы организма в состоянии покоя, в том числе и при поворотах головы и движениях рук, а в положении сидя ног, вызывающих смещение центра тяжести. К ним относятся сегментарные и надсегментарные (шейные) тонические рефлексы спинного мозга и рефлексы ствола мозга, возникающие с вестибулорецепторов. Их роль – сохранение естественной позы. Для выявления роли шейных рефлексов Магнус удалял у животного вестибулярный аппарат, чтобы погасить вестибулярные рефлексы. Для выявления вестибулярных рефлексов на шею животного, расположенную на одной линии с туловищем, накладывают гипсовую повязку, чтобы прекратить шейные надсегментарные рефлексы. Вестибулярные рефлексы выявляются при поворачивании животного через ось висков. Если животное располагается горизонтально спиной и затылком вниз и до 45 градусов выше горизонтали, то тонус мышц разгибателей становится максимальным; если при дальнейшем повороте оно располагается спиной и затылеом вверх по горизонтали и до -45 градусов по вертикали, то тонус мышц разгибателей становится максимальным. Статокинетические рефлексы – рефлекторное перераспределение мышечного тонуса, возникающее с кожных, проприо и вестибулорецепторов, при нарушении позы. К ним относятся выпрямительные рефлексы – выпрямление головы и туловища. Выпрямление головы запускается с вестибулярных и кожных рецепторов, выпрямление туловища с проприорецепторов шеи и кожных рецепторов.
РФ образована нейронами, расположенными диффузно и в виде ядер в центральных отделах ствола мозга. Они имеют хорошо ветвящиеся аксоны, которые позволяют одному нейрону устанавливать связи с 25 тыс. других нейронов; принимают импульсы от сенсорных путей, идущих от разных рецепторов. Ретикулярные нейроны полимодальные; имеют большие рецепторные поля; у них длительные латентный период ответа на периферическую стимуляцию, связанный с проведением возбуждения до них через многочисленные синапсы. Они имеют фоновую тоническую активность, высокие возбудимость и мобильность, чувствительны к некоторым веществам крови (адреналин, СО2) и лекарствам (барбитураты, аминазин) РФ имеет афферентные входы преимущественно от трех источников: 1. От температурных и болевых рецепторов по волокнам спиноретикулярного пути тройничного нерва; 2. От сенсорной и частично других зон коры головного мозга по кортикоретикулярным путям импульсация поступает в ядра, дающие начало ретикулярным путям, а также ядрам, которые проецируются на мозжечок; 3. От ядер мозжечка по мозжечковоретикулярному пути. Эфферентные выходы: 1. В спинной мозг – по медиальному и латеральному ретикулоспинальным путям; 2. К верхним отделам головного мозга (неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу) идут восходящие пути, начинающиеся в ядрах продолговатого мозга и моста; 3. К мозжечку идут пути, которые начинаются в латеральном и парамедианном ретикулярном ядрах и я ядре покрышки моста. Восходящее влияние на большой мозг. РФ может оказывать как активирующее, так и тормозное действие. Оно образуется аксонами 5 – 17 % нейронов РФ, формирующими ретикулоталамические пути. Их стимуляция приводит к появлению постсинаптических потенциалов в нейронах и вызванных потенциалов в неспецифических и специфических ядрах таламуса. После переключения в нейронах таламуса сенсорный поток проецируется в различные области коры. Восходящие влияния поступают в задний таламус, полосатое тело, прозрачную перегородку. В восходящей РФ выделяют две части: ростральную, образуемую нейронами РФ среднего мозга, и каудальную, формируемую нейронами РФ продолговатого мозга и моста. Активирующее влияние ростральной части РФ впервые было показано с помощью перерезки ствола мозга между верхним и нижним холмиками четверохолмия. В результате перерезки животное погружалось в сноподобное состояние: нарушен контакт с внешним миром, не реагирует на световые и обонятельные раздражители. Прямое доказательство активирующего влияния РФ на большие полушарие было получено в опытах с раздражением РФ через погружные электроды у спящих животных (Мегун, Моруцци). Стимуляция РФ вызывала пробуждение животного. У него возникала ориентировочная реакция, в корковых нейронах возникали ВПСП; альфа ритм и более медленные ритм сменялись бета ритмом – реакция десинхронизации ЭЭГ. Активирующее влияние на кору осуществляется в основном при помощи холинергических нейронов. Тормозное влияние возникало при стимуляции каудального отдела РФ. При этом на корковых нейронах формировались в основном ТПСП и выявлялся синхронизирующий эффект на ЭЭГ. Тормозное влияние вызывает синхронизацию ЭЭГ в фазе медленного сна и атонию мышц в фазе быстрого сна. У бодрствующих животных стимуляция каудальной РФ угнетает локомоторную активность, но существенно не влияет на рефлексы пищевого поведения. Тормозные влияния осуществляются также при помощи холинергических нейронов.
Нисходящие влияние на тонус мышц и формирование позы РФ оказывает через латеральный и медиальный ретикулоспинальные тракты. Медиальный ретикулоспинальный путь идет от ретикулярных ядер моста и возбуждает спинальные альфа и гамма нейроны мышц – разгибателей осевой мускулатуры (туловища, проксимальных отделов конечностей), через тормозные интернейроны тормозятся мотонейроны сгибателей. Латеральный ретикулоспинальный путь идет от ретикулярного гигантоклеточного ядра продолговатого мозга, возбуждаются альфа и гамма нейроны мышц сгибателей мускулатуры конечностей и реципрокно затормаживают мышцы – разгибатели. Взаимоотношения между стволовыми и двигательными центрами выявляются в эксперименте с перерезкой ствола по передней границе четверохолмия. У подопытных животных в этом случае повышается тонус мышц разгибателей конечностей, туловища, шеи, хвоста – децеребрационная ригидность. 
Выделяют три структуры мозжечка. Древний мозжечок состоит из клочка и узелка, а также часть червя. Старый мозжечок включает центральную дольку, вершину пирамиду, язычок червя, и четырехугольную дольку полушарий. Новый мозжечок состоит из полушарий и части червя (скат, листок, бугор). Клетки Пуркинье, образующие средний слой коры, являются главной функциональной единицей с ярко выраженной интегративной направленностью. Она реализуется сильно ветвящимися дендритами, на которых в одной клетке может быть до 100 тыс синапсов. Клетки Пуркинье являются единственными эфферентными нейронами коры мозжечка, связывая ее с внутримозжечковыми и вестибулярными ядрами. К коре мозжечка проходит три вида волокон – лиановые, моховидные и моноаминергические. Лиановые волокна являются аксонами нейронов нижней оливы продолговатого мозга. Инфа к ним поступает от мышечных, кожных рецепторов и двигательной коры головного мозга. Каждое лиановидное волокно устанавливает контакт с дендритами одной клеткой Пуркинье, но число синапсов может достигать 300, что объясняет исключительно сильное возбуждающее действие этого афферентного входа. Также они оказывают тормозящие действие на клетки Пуркинье через корзинчатые и звездчатые клетки поверхностного слоя мозжечка. По моховидным волокнам в кору мозжечка поступает инфа от коры больших полушарий , проприорецепторов, вестибулярных рецепторов и РФ. Они образуют синапсы на дендритах клеток зерен внутреннего слоя коры. Через аксоны клеток – зерен моховидные волокна оказывают возбуждающее влияние на клетки Пуркинье и опосредованно тормозное действие через корзинчатые и звездчатые клетки. Активность клеток – зерен регулируется через тормозные клетки Гольджи внутреннего слоя по типу возвратного торможения (медиатор ГАМК). Монаминергический афферентный вход. Через него на мозжечок передается влияние голубого пятна (медиатор норадреналин) и ядер шва (серотонин). Стимуляция этих ядер тормозит фоновую активность клеток Пуркинье и модулирует их ответы на активацию моховидных и лиановидных афферентов. На клетки Пуркинье поступает в основном возбуждающее влияние. Но их возбуждение оказывает тормозное влияние на три парных мозжечковых ядра (ядра шатра, зубчатые и промежуточные), иннервирующих моторные центры головного мозга. Мозжечок осуществляет регуляцию по механизму сдерживания активности моторных центров. Три парных ядра мозжечка образуют три главных эфферентных выхода на стволовые и корковые моторные центры. Нейроны этих ядер имеют высокую фоновую активность. Они находятся под тормозящим влиянием клеток Пуркинье и возбуждающим влиянием афферентных входов в мозжечок, которые по коллатеральным ответвлениям поступают в эти ядра. Двигательные функции мозжечка связаны с координацией движений и обучением двигательным навыкам. Регуляция мышечного тонуса, обеспечивающего поддержание естественной позы (равновесия) осуществляется преимущественно древним и частично старым мозжечком. Получая и обрабатывая импульсацию от вестибулярных рецепторов, проприорецепторов аппарата движения, кожных, зрительных и слуховых рецепторов, мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, а также прямого выхода на вестибулярные ядра изменить позу тела и сохранить равновесие. Мозжечок осуществляет тонкую настройку вестибулярных рефлексов, в том числе рефлекторное поддержание вертикальной позы. Координация движений осуществляется старым и новым мозжечком. В кору этой части мозжечка поступает инфа от рецепторов аппарата движения, а также импульсация от моторной коры. Анализируя инфу о программе и выполнении движения, мозжечок способен через свои промежуточные ядра, имеющие выходы на красное ядро и моторную кору, осуществить координацию, а при необходимости и коррекцию движений. При этом регулируются амплитуда, направление и темп движения. Программирование произвольных движений осуществляется корой нового мозжечка, которая получает инфу о замысле движения из ассоциативных зо коры большого мозга через ядра моста. В коре нового мозжечка она перерабатывается в программу движения, которая через зубчатое ядро мозжечка и вентральное латеральное ядро таламуса поступает в премоторную зону. Там она получает дальнейшую обработку и через пирамидную и экстрапирамидную системы реализуется как сложное целенаправленное движение. Контроль коррекция и координация медленных програмированных движений осуществляется на основе обратной афферентации от проприорецепторов, а быстрых путем изменения программы движения без сличения с обратной афферентацией. |