Физиология нервов, синапсов, мышц и рецепторов
 Скачать 23.71 Mb. Скачать 23.71 Mb.
|

|
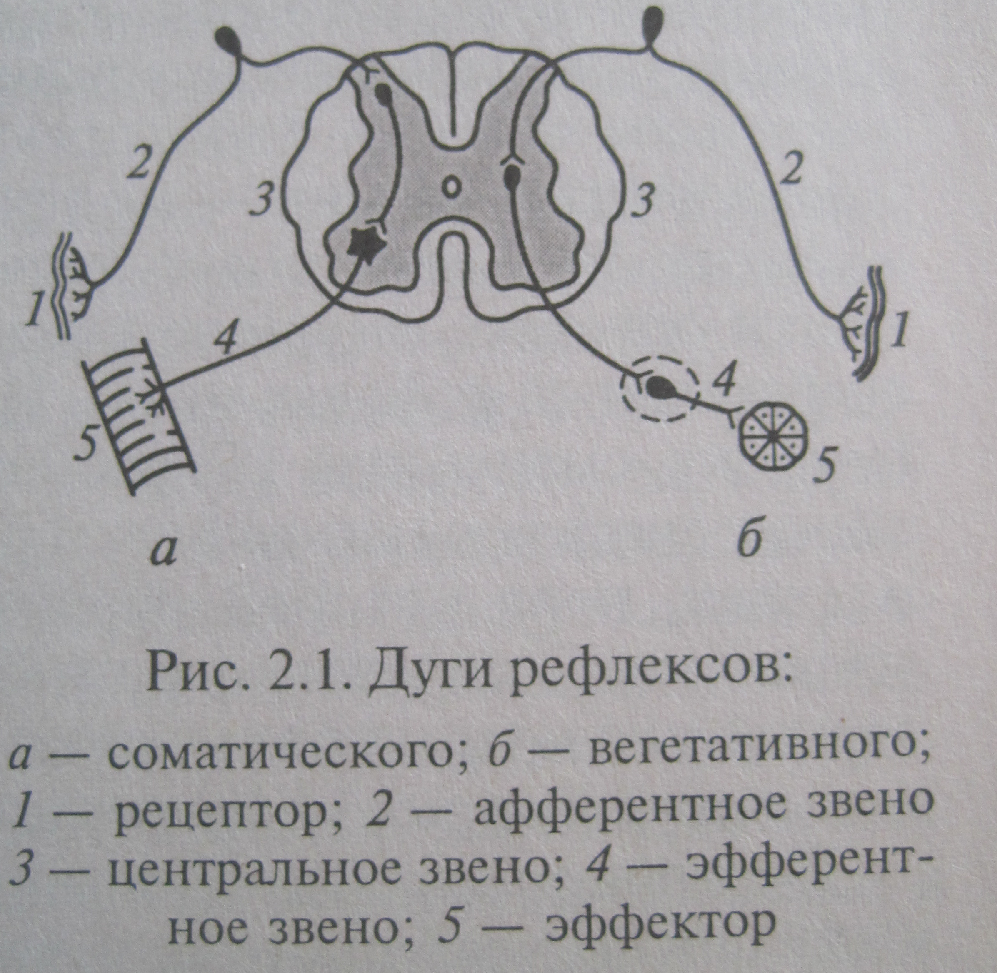
Типы регуляции. Рефлекс. Универсальность и приспособленный характер изменчивости рефлекса, развитие концепции, рефлекс (Декарт, И.М.Сеченов, И.П.Павлов, П.К.Анохин). Рефлекс – ответная реакция организма на раздражение сенсорных рецепторов, осуществляемая с помощью НС. Каждый рефлекс осуществляется посредством рефлекторной дуги, которая состоит из следующих структур.
Рефлексы классифицируют по срокам появления в онтогенезе: врожденные и приобретенные. Безусловные делят на несколько групп:
Развитие концепции рефлекса. На первом этапе были сформулированы основные положения о принципе деятельности ЦНС французским естествоиспытателем Декартом. Он сформулировал два важных положения рефлекторной теории: 1. Реакция организма на внешние воздействия является отраженной. 2. Ответная реакция на раздражение осуществляется при помощи НС. На втором этапе было экспериментально обосновано материалистическое представление о рефлексе. Было установлено, что рефлекторная реакция может осуществляться на одном метамере лягушки. Выявлено, что стимулы могут быть не только внешними, но и внутренними; установлена роль задних чувствительных и передних двигательных корешков спинного мозга. Сеченов обосновал приспособительный характер изменчивости рефлекса, открыл центральное торможение, а также явление суммации возбуждения в ЦНС. На третьем этапе были открыты условные рефлексы и разработаны основы учения о ВНД. Павлов сформулировал три принципа рефлекторной теории: 1. Принцип детерминизма, согласно которому любая рефлекторная реакция причинно обусловлена – возникает при действии раздражителя; 2. Принцип структурности, суть которого заключается в том, что каждая рефлекторная реакция осуществляется при помощи определенных структур, и чем больше структурных элементов участвует в осуществлении этой реакции, тем она совершеннее; 3. Принцип единства процессов анализа и синтеза – НС анализирует с помощью рецепторов все действующие внешние и внутренние раздражители и на основании этого анализа формирует целостную ответную реакцию (синтез). На четвертом этапе было создано учение о функциональных системах (Анохин).
Классификация нейронов. По обеспечению соматических или висцеральных функций подразделяются на соматические и вегетативные соответственно двум отделам нервной системы. По месту в системе рефлекторных связей (соответственно по направлению или источнику информации) на афферентные, воспринимающие с помощью сенсорных рецепторов инфу о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; эфферентные, передающие инфу к рабочим органам – эффекторам; вставочные, обеспечивающие взаимодействие между нейронами ЦНС. По хим природе медиатора, выделяющегося в окончаниях аксонов, адренергические, холинергические, серотонинергические и др. По типу ответной реакции эффектора тормозящие и возбуждающие. Нейроны высших отделов ЦНС по специфичности воспринимаемой сенсорной информации могут быть моно, би и полисенсорными. По характеру активности фоноактивные и молчащие, возбуждающимися только в ответ на раздражение. Фоноактивные нейроны играют большую роль в поддержании тонуса ЦНС и особенно коры большого мозга. Классификация синапсов. В зависимости от местоположения аксо – соматические, аксодендритные, аксоаксонные, сома – соматические, дендросоматические, дендродендритные. По рабочему эффекту возбуждающие и тормозящие. По способу передачи сигналов химические, электрические, смешанные. Свойства химических синапсов. Большинство синапсов являются химическими. Первое их преимущество заключается в том, что хим синапсы благодаря наличию различных медиаторов и постсинаптических мембран могут оказывать возбуждающие и тормозное влияние на нервные и эффекторные клетки. Второе – могут передавать инфу в широких временных рамках. Третье – высокая пластичность; их функция зависит от предшествующей нейрональной активности, поэтому они играют важную роль в процессах обучения и памяти. Химические синапсы. На пресинаптической поверхности активной зоны сос торны синаптической везикулы находятся филаменты и частицы, выступающие из пресинаптической терминали, часто формирующие правильный шестиугольник, названный пресинаптической решеткой. Определенные точки на сетке являются участками экзоцитоза синаптических везикул. Медиатор в пресинаптическом окончании находится в двух пулах – готовом к экзоцитозу (пул 1 – медиатор лежит на пресинаптической мембране изнутри) и мобилизационном (пул 2 – находится в цитоскелете), из которого пополняется в течение нескольких секунд пул 1. Этоо процесс активируется ионами кальция. Химическим синапсам свойственно явление суммации. Электросекреторное сопряжение в ЦНС обеспечивается ионами кальция: при возбуждении пресинаптической терминали открываются потенциалзависимые кальциевые каналы. Кальций входит в нервное окончание согласно электрохимическому градиенту и взаимодействует с экзоцитозным аппаратом пресинапса, представляющим собой совокупность белков. Активация белков кальцием вызывает выброс медиатора в синаптическую щель. Синаптическая щель хим синапсов содержит поперечно расположенные гликопротеиновые филаменты, обеспечивающие связи пре и постсинаптической мембран и направление диффундирующего медиатора. Медиатор взаимодействует с белком – рецептором постсинаптической мембраны, и эффекторная клетка приходит в состояние возбуждения или торможения. Электрические возбуждающие синапсы. Обнаружены в составе мезенцефального ядра тройничного нерва, вестибулярного ядра Дейтерса, ядра нижней оливы продолговатого мозга. Характерны для сердца, гладких мышц, секреторных клеток. Имеются следующие электрические синапсы: аксон – сома, аксон – дендрит, аксон – аксон, дендрит – дендрит, дендрит – сома, сома – сома. Проводят сигнал в обе стороны без синаптической задержки, передача не блокируется при удалении кальция. Контактирующие мембраны клеток в составе электрического синапса связаны друг с другом полуканалами белковой природы – коннексонами. Через коннексоны клетки обмениваются аминокислотами, пептидами, РНК, метаболитами, циклическими нуклеотидами. Механизм передачи возбуждения в электрическом синапсе: ПД, возникающий на мембране одной клетки, с помощью своего электрического поля деполяризует мембрану соседней клетки до КП.
Механизм возбуждения нейронов.
Поступившей в пресинаптическое окончание кальций удаляется из него при помощи кальциевого насоса. Прекращение действия медиатора осуществляется посредством обратного захвата пресинаптическим аппаратом, с помощью разрушения специальными ферментами. Норадреналин разрушается монаминоксидазой и катехолметилтрансферазой, АХ гидролизуется ацетилхолинтрансферазой. Прекращение действия избытка медиатора на постсинаптическую мембрану предотвращает десенситизацию – снижение чувствительности постсинаптичекой мембраны к действующему медиатору. Пептидные медиаторы, ферменты и другие белки транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофиламентов, тянущихся по всей длине аксона. Для этого транспорта необходимы кальций и энергия.
Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах от аксона одного нейрона к телу или дендритам другого объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении, т.к. пресинаптическая мембрана чувствительна только к электрическому импульсу, а постсинаптическая к медиатору. Замедленное распространение возбуждения в ЦНС по сравнению с нервным волокном объясняется наличием на путях распространения возбуждения многих химических синапсов, в каждом из которых имеется синаптическая задержка около 0,5 мс. Суммарная задержка передачи возбуждения достигает 2 мс. Дивергенция возбуждения в ЦНС объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенцию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное - энергичное движение всех конечностей или даже туловища. Конвергенция возбуждения (принцип общего конечного пути) – схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип воронки Шеррингтона). Объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов. Циркуляция возбуждения по замкнутым нейронным цепям может продолжаться несколькими минутами или даже часами. Циркуляция возбуждения – одна из причин явления последействия. Пресинаптическое усиление: в аксо – аксональных возбуждающих синапсах возбуждение одного нервного окончания обеспечивает деполяризацию другого нервного окончания. Вследствие этого в деполяризированное окончание входит кальций, который способствует выделению медиатора Распространение возбуждения в ЦНС легко блокируется фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Свойства нервных центров.
Кратковременная после одиночных стимулов выражена слабо, после раздражающей серии потенциация в ЦНС длится дольше. Одной из причин возникновения является накопление кальция в пресинаптических окончаниях. Соответственно количество медиатора, выделяемого при каждом стимуле, увеличивается, возрастает ВПСП. Долговременная потенциация – быстро развивающееся устойчивое повышение возбудимости постсинаптического нейрона при высокочастотных повторных или кратковременных сильных активациях пресинаптического нейрона. Связана с внутриклеточным повышением концентрации ионов кальция, вызванной активацией постсинаптических nmda – рецепторов глутаматом. Эти рецепторы являются кальциевыми каналами, пропускающими в клетку большие количества ионов кальция и натрия. В неработающем организме эти рецепторы закрыты магнием. Сильное возбуждение выбивает магниевую пробку. Вошедшие в клетку натрий и кальций вызывают значительную деполяризацию клетки, приближающуюся к КП. В этих условиях достаточно даже миниатюрных ВПСП, чтобы возникло возбуждение постсинаптического нейрона. Значение синаптической потенциации заключается в том, что она создает предпосылки улучшения процессов переработки информации в нейронах нервных центров. Может вызвать переход центра в доминантное состояние. Доминанта – стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Открыл Ухтомский в опытах с раздражением двигательных зон коры большого мозга и наблюдением сгибания конечности животного. Если раздражать корковую двигательную зону на фоне сильного повышения возбудимости другого нервного центра, обычного сгибания конечности не происходит. Доминантный очаг возбуждения обладает следующими свойствами: стойкость, повышенная возбудимость, способность притягивать к себе иррадиирущие по ЦНС возбуждения и оказывать угнетающее воздействие на центры конкуренты и др нервные центры. Значение доминантного очага возбуждения состоит в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии. Синаптическая депрессия (утомляемость нервных центров) – ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапсов. Объясняется расходованием медиатора, накоплением метаболитов, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям. Компенсация нарушенных функций после повреждения того или иного центра – результат проявления пластичности ЦНС. Нарушенная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и вовлечения в эту функцию других рассеянных нейронов в коре большого мозга с подобными функциями. Адаптация к протезам является проявлением пластичности. Существуют следующие методы исследования функций ЦНС: 1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом. 2. Метод экстирпации (удаления) или разрушения участков мозга. 3. Метод раздражения различных отделов и центров мозга. 4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием. 5. Электрофизиологические методы: а. электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г. Бергером. б. регистрация биопотенциалов различных нервных центров; используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро. в. метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков; 6. метод внутримозгового введения веществ с помощью микроинофореза; 7. хронорефлексометрия – определение времени рефлексов. |