Глик Молекулярная биотехнология. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. Пер с англ. М. Мир, 2002. 589 с
 Скачать 9.74 Mb. Скачать 9.74 Mb.
|

ГЛАВА 4. | ||||||||||||||||||||||||||||||||||||||||
| Рис. 4.1. Клонирование рекомбинантной ДНК. Донорную ДНК расщепляют рестрицирующей эндонуклеазой и встраивают в клонирующий вектор. Полученную конструкцию вводят в популяцию клеток-хозяев, идентифицируют те клетки, которые содержат рекомбинантную ДНК, и культивируют их. При необходимости можно индуцировать экспрессию клонированного гена в клетках-хозяевах и получить кодируемый белок | 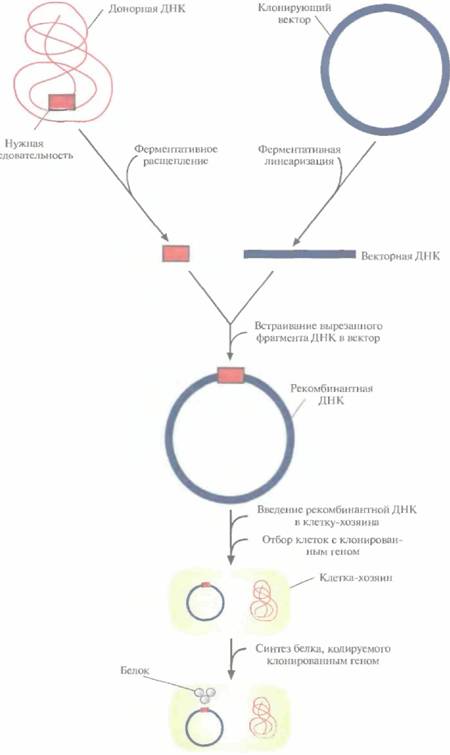 |
52 ГЛАВА 4
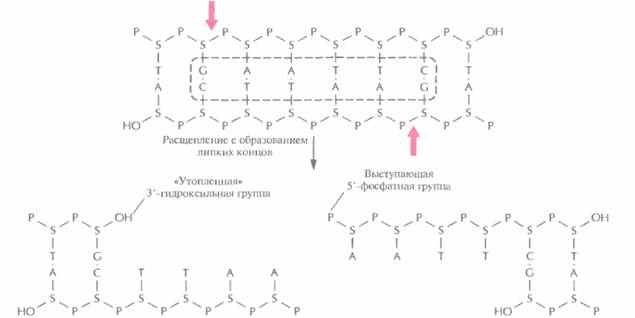 | |
| Риc. 4.2. Расщепление короткого фрагмента ДНК рестрицирующей эндонуклеазой типа II EcoRI с образованием липких концов. Стрелки — связи, по которым происходит расшепление в сахарофосфатном остове. S — деэоксирибоза, P — фосфатная группа, ОН — гидроксильная группа. Последовательность, распознаваемая EcoRI, выделена штриховой линией. | |
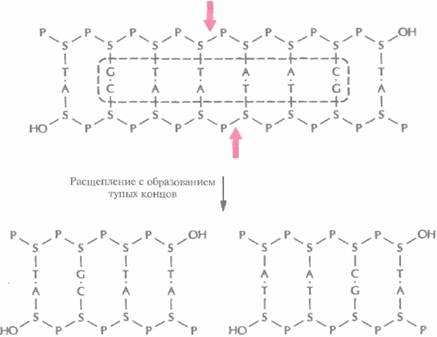 | |
| Рис. 4.3. Расщепление короткого фрагмента ДНК рестриктазой типа II HindIIс образованием тупых концов. Стрелки — связи, по которым происходит расщепление в сахарофосфатном остове. Буквенные обозначения - те же, что и на рис, 4.2. Последовательность, распознаваемая рестрикгазой HindII, выделена штриховой линией. | |
Технология рекомбинантных ДНК 53
и получила назваие EcoRI. Этот фермент узнает участок ДНК, содержащий специфическую палиндромную последовательность (последовательность-перевертыш, идентичную в обеих цепях при прочтении в направлении 5'-->3') из шести пар оснований и вносит разрыв между остатками гуанина и аденина в каждой цепи (рис. 4.2), расщепляя связь между атомом кислорода при 3'-атоме углерода сахарного остатка одного нуклеотида и фосфатной группой, присоединенной к 5 '-углеродному атому сахарного остатка соседнего нуклеотида. Разрывы в цепи ДНК располагаются наискось друг от друга, в результате чего образуются одноцепочечные комплементарные концы с «хвостами» из четырех нуклеотидов в каждом (липкие концы). Каждый одноцепочечный «хвост» заканчивается 5'-фосфатной группой, а 3'-гидроксильная группа противоположной цепи как бы утоплена.
Помимо EcoRI, из бактериальных клеток были получены сотни рестрицирующих эндуклеаз типа II. Названия этим эндонуклеазам даются по такому же принципу, как и .EcoRI: род микроорганизма обозначается прописной буквой, а вид — двумя строчными; штамм обычно не указывается. Римские цифры — порядковый номер данной эндонуклеазы в ряду прочих рестриктаз, выделенных из данного микроорганизма. Например, HраIи НраII — это соответственно первая и вторая рестрицируюшие эндонуклеазы типа II, выделенные из Haemophilus parainfluenzae.
Палиндромные последовательности, которые распознаются рестрицирующими эндонуклеазами типа II и в которых происходит расщепление молекулы ДНК, называются сайтами узнавания. Помимо рестриктаз, гидролизующих (расщепляющих) полинуклеотидную цепь с образованием липких концов, существуют рестриктазы, которые вносят разрывы в цепи строго друг против друга с образованием фрагментов ДНК с «тупыми» концами (рис. 4.3). Сайты узнавания могут состоять из четырех, пяти, шести, восьми или более пар нуклеотидов (табл. 4.1). От длины сайта узнавания зависит частота его распространения в молекуле ДНК; в большинстве случаев используют рестриктазы, узнающие тетра- и гексануклеотиды.
Рестрицирующие эндонуклеазы типа II играют ключевую роль при генном клонировании.
| Таблица 4.1. Нуклеотидные последовательности, распознаваемые некоторыми ферментами рестрикции | ||
| Фермент | Сайт узнавания | Характер образуемых концов |
| | | |
| | | |
| | | |
| | | |
| | | |
| | | |
| | | |
| | | |