Физа. экзамен физа. Итоговое по разделу Общая физиология Теоретические вопросы
 Скачать 0.79 Mb. Скачать 0.79 Mb.
|

|
Название рецептора определено медиатором, с которым он взаимодействует: холинорецепторы, адренорецепторы, дофаминовые рецепторы, серотониновые /триптаминовые/ рецепторы, гистаминовые рецепторы, ГАМК-рецепторы, эндорфиновые рецепторы и т.д. Медиаторы обладают 2 видами действия 1.ионотропное - изменяют проницаемость каналов для ионов 2.метаботропное - через вторичные посредники запускают и тормозят соответствующие процессы в клетках. Медиаторы - это биологически активные вещества, синтезируются в нервных клетках. Они концентрируются и выделяются только в месте контакта нейрона с другими клетками. Все медиаторы можно разделить на возбуждающие медиаторы и тормозные медиаторы. И синапсы делятся на возбуждающие и тормозные. Возбуждающие медиаторы взаимодействуя с рецептором субсинаптической мембраны вызывают активацию натриевых каналов и формируют входящий натриевый ток, который вызывает возникновение частичной деполяризации, т.е рецепторный потенциал, который на уровне синапса обозначают как возбуждающий постсинаптический потенциал (ВПСП). Тормозной медиатор вызывает усиление входящего калиевого тока или входящего тока хлора, т.е. вызывает локальную гиперполяризацию. Это формирует тормозной постсинаптический потенциал (ТПСП). Конечный эффект (потенциал действия или тормозной потенциал) формируется за счет суммации ВПСП или ТПСП. Инактивация медиатора: В естественных условиях медиатор отсоединяется от рецепторов и разрушается ферментами (холинэстераза и т.д.), которые имеются в синапсе. Примерно 20-30% медиатора удаляется таким образом из синаптической щели – первый способ инактивации. Инактивация медиатора Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание. Аптейк - обратный захват пресинаптической мембраной. 7. Торможение в ЦНС (И.М. Сеченов). Современная классификация, клеточные механизмы центрального торможения. Тормозящие синапсы и их медиаторы. Тормозной постсинаптический потенциал (ТПСП). Виды центрального торможения в нервных сетях. Принципы координации (упорядочения) рефлекторной деятельности. В 1862 г. И. М. Сеченов открыл явление центрального торможения. Он доказал в своем опыте, что раздражение кристалликом хлорида натрия зрительных бугров лягушки (большие полушария головного мозга удалены) вызывает торможение рефлексов спинного мозга. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась. Результат этого опыта позволил И. М. Сеченому сделать заключение, что в ЦНС наряду с процессом возбуждения развивается процесс торможения, который способен угнетать рефлекторные акты организма. Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную формацию ствола мозга): возбуждение ретикулярной формации повышает активность тормозных нейронов спинного мозга – клеток Реншоу, что приводит к торможению α-мотонейронов спинного мозга и угнетает рефлекторную деятельность спинного мозга. Торможение– активный процесс, возникающий при действии раздражителей на ткань, проявляется в подавлении другого возбуждения. Выделяют два типа торможения: 1) первичное. Для его возникновения необходимо наличие специальных тормозных нейронов. Возникает под воздействием тормозного медиатора. Различают два вида первичного торможения: - пресинаптическое в аксо-аксональном синапсе; - постсинаптическое в аксо-дендрическом синапсе. 2) вторичное. Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур Виды вторичного торможения: - запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона; - пессимальное, возникающее при высокой частоте раздражения; парабиотическое, возникающее при сильно и длительно действующем раздражении; Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. -65мВ до -70мВ, следовательно, проведение нервного сигнала через синапс тормозит ТПСП, равный -5 мВ. 8. Вегетативная нервная система. Морфофункциональная характеристика и особенности функционирования вегетативной нервной системы по сравнению с соматической. Сравнительная морфофункциональная характеристика симпатического и парасимпатического отделов вегетативной нервной системы. Вегетативные рефлексы, их классификация и характеристика. Вегетативная нервная система – эфферентный отдел нервной системы, обеспечивающий регуляцию деятельности внутренних органов. 1. В отличие от Соматической нервной системы Вегетативная обладает выраженной автономией (не только от коры Больших Полушарий, но зачастую и от ЦНС в целом.) 2. Двухнейронность - содержит два класса нейронов (1-й и 2-й нейроны ВегНС). 3. Низкая возбудимость нейронов ВНС 4. Наличие ганглиев - часть ЦНС, вынесенная в ходе эволюции на периферию. 5. Наличие двух типов нервных волокон: Двухнейронность и наличие ганглия позволяет выделить преганглионарное волокно и постганглионарное.. Наличие медленного преганглионарного волокна (представленного типом В), ещё более медленного постганглионарного волокна (типа С) и дополнительного синапса с синаптической задержкой обуславливают более низкую скорость проведения возбуждения. 6.Двухнейронность подразумевает и наличие двух синапсов (центральный и периферический.) 7. ВНС представлена двумя отделами: СНС и ПСНС 8. Очаговое представительство нервных центров СНС и ПСНС в ЦНС. Тела первых нейронов располагаются: 1. СНС - боковые рога торако-люмбального отдела спинного мозга. 2. ПСНС - три зоны, где лежат её центры: а) мезенцефальный отдел (ветви в составе глазодвигательного нерва - зрачок, некоторые слюнные железы); б) бульбарный отдел - лицевой, языкоглоточный нерв и n . vagus ; в) сакральный отдел - центры ПС иннервации органов малого таза. Отличия в строении и функционировании отделов ВНС: 1. Центры: - СНС - расположены компактно, пространственно объединены. - ПСНС - резко-выраженная очаговость. 2. Ганглии: - СНС - близко к позвоночному столбу; формируют пара- и превертебральные цепочки. ПСНС - рядом с органом-исполнителем или интрамурально, не связаны между собой. 3. Нервные волокна: - СНС - преганглионарные волокна короткие, постганглионарные - длинные. - ПСНС - преганглионарные волокна - длинные, постганглионарные - короткие. 4. Реакция: - СНС – генерализованная. - ПСНС - локальная, регионарная. Функции Вегетативной нервной системы: 1. Триггерное влияние - явление запуска функции какого-либо органа. 2. Корригирующее влияние - т.к. ряд органов обладает автоматией, то активация того или иного отдела Вегетативной нервной системы приведет лишь к изменению активности органа. 3. Адаптационно-трофическое влияние (характерно для симпатической нервной системы). СНС не только непосредственно влияет на функцию, но и на процессы питания тканей и интенсивность обмена веществ. Вегетативные рефлексы Рефлекторные дуги соматических и вегетативных рефлексов отличаются лишь эфферентной частью. Поэтому любое воздействие, как правило, сопровождается как реакцией со стороны скелетной мускулатуры, так и со стороны внутренних органов (т.е. можно выделить в реакции организма двигательный и вегетативный компонент). 1. Висцеро-висцеральные - (изменение АД - изменение работы сердца). 2. Висцеро-кутанные - при заболевании внутренних органов - изменения чувствительности, парэстезии,изменение потоотделения. 3. Кутано-висцеральные- горчичники, банки, массаж, иглорефлексотерапия. 4. Висцеро-соматические- раздражение хеморецепторов каротидного синуса СО2 стимулирует работу дыхательной мускулатуры (межреберные мышцы). 5. Сомато-висцеральные - физ. работа приводит к изменению деятельности ССС и системы дыхания. Примеры вегетативных рефлексов, наиболее часто оцениваемых в практической медицине: 1. Болевой рефлекс- активирует СНС (диагностика чувствительности). 2. Рефлекс Гольца- раздражение петель кишечника, брюшины приводит к урежению или остановке сердцебиений (активация ПСНС). 3. Рефлекс Данини-Ашнера- (глазо-сердечный рефлекс) -надавливание на глазные яблоки (повышение внутриглазного давления) - урежение сердечных сокращений (применяется для снятия пароксизмальной тахикардии). 4. Рефлекс Ортнера - запрокидывание головы - изменение параметров работы сердца. 5. Дыхательная аритмия (дыхательно-сердечный рефлекс) - урежение сердцебиений в конце выдоха. 6. Ортостатическая реакция - повышение ЧСС и АД при переходе из горизонтального в вертикальное положение. 7. Дермографизм - в норме - белый, при повышенной чувствительности - красный. 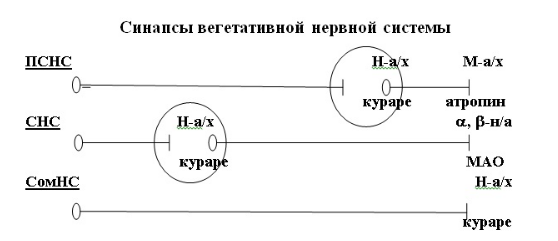 Начало формы Начало формыНачало формы 9. Парасимпатическая нервная система, морфофункциональная характеристика, влияние на функцию органов и систем организма. Холинэргические синапсы. Система образования и инактивации ацетилхолина. Понятие о М- и Н- холинорецепторах. Ацетилхолин - медиатор ганглионарных синапсов симпатической и парасимпатической нервной системы, периферического синапса парасимпатической нервной системы и нервно-мышечного синапса соматической нервной системы. Разрушается холинэстеразой. Холинэргические синапсы — синапсы, в которых передача возбуждения осуществляется посредством ацетилхолина. Нервный импульс провоцирует высвобождение молекул ацетилхолина в синаптическую щель. В парасимпатической НС медиатором является ацетилхолин, рецепторами – холинорецепторы двух типов: Н (никотин) и М (мускарин). М-холинорецепторы - чувствительны к мускалину (яду мухомора) - расположены в основном во внутренних органах, эндокринных железах, сердце, сосудах, дыхательных путях, желудочнокишечном тракте. Они обладают медленным, но продолжительным действием, могут суммировать возбуждение. Существуют 2 вида М-холинорецепторов: одна - во внутренних органах, другая - в эндокринных железах. При возбуждении М-холинорецепторв происходит торможение сердечной деятельности, расширение сосудов, активация деятельности желудочно-кишечного тракта, изменяется секреция некоторых эндокринных желёз. Н-холинорецепторы - чувствительны к никотину. Располагаются в вегетативных ганглиях, мионевральных синапсах, в хлорофильной ткани надпочечников. Эти рецепторы обладают быстрым, кратковременным действием, не могут суммировать возбуждение. Существует 3 разновидности. В ЦНС больше Н-холинорецепторов. М-холинорецепторы преобладают в области ствола мозга, подкорковых узлах, лимбической системе, ретикулярной формации, гипоталамусе. 10. Симпатическая нервная система, морфофункциональная характеристика, влияние на функцию органов и систем организма. Холинэргические синапсы в ганглиях. Адренэргические синапсы. Система образования и инактивации катехоламинов. Понятие об альфа- и бета-адренорецепторах. Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина. Адренергические синапсы располагаются в органах, получающих постганглионарную симпатическую иннервацию. Синапс представляет собой контакт окончания адренергического нерва с клеткой рабочего органа. Передача импульса через адренергический синапс осуществляется с помощью медиатора норадреналина. Инактивация катехоламинов происходит двумя путями: 1) в результате дезаминирования под действием фермента моноаминоксидазы (МАО) 2) метилирования под действием фермента катехол-О-метилтрансферазы (КОМТ). Норадреналин – медиатор периферического синапса симпатической нервной системы (адренэргический синапс). Кроме норадреналина, в качестве медиатора, могут быть использованы адреналин и дофамин. Синтезируется норадреналин из аминокислоты тирозина. Разрушается МАО (моноаминоксидазой) после обратного захвата из синаптической щели и КОМТ (катехоламинтрансферазой) в области рецепторов. Рецепторы постсинаптической мембраны вегетативных синапсов. В ганглионарных синапсах симпатической и парасимпатической нервной системы передача возбуждения блокируется никотином (Н-холинорецепторы). В периферическом синапсе парасимпатической нервной системы передача возбуждения блокируется мускарином (М-холинорецепторы). В периферических синапсах симпатической нервной системы адренорецепторов неодинаковы (альфа и бета – адренорецепторы). Воздействие медиатора на альфа- адренорецепторы вызывает сокращение гладкой мускулатуры, а на бета - адренорецепторы – расслабление гладкой мускулстуры. 11. Гуморальная регуляция функций, характеристика, место и роль в регуляции функций. Уровни регуляции. Классификация БАВ. Местная регуляция, механизмы ее реализации. Креаторные связи, роль неспецифических метаболитов. Региональный уровень регуляции, роль специфических и неспецифических метаболитов, функциональная характеристика основных парагормонов. I уровень: местная или локальная регуляция происходит на минимальном пространстве, касается ограниченного числа клеток (единицы, десятки). 1. креаторные связи и кейлоны Креаторные связи - обмен между клетками макромолекулами, несущими информацию о клеточных процессах, позволяющую клеткам ткани функционировать содружественно. Это один из наиболее эволюционно старых способов регуляции. Кейлоны - вещества, обеспечивающие креаторные связи. Обеспечивается только гуморальными факторами: - в основном НМ (неспецифическими метаболитами) и -в меньшей степени - СМ (специфическими метаболитами). II уровень регуляции: региональный (органный). Регуляция осуществляется в пределах органа, сосудистого региона. Центральное место в этой регуляции играют органные ганглии III уровень - межорганное, межсистемное регулирование. 1. Гуморальная регуляция представлена железами внутренней секреции. б) истинные гормоны вырабатываются в железах внутр секреции (ЖВС) Истинные гормоны - БАВ, вырабатывающиеся в специализированных железах внутренней секреции, обладающие дистантным действием и высокой активностью. Делятся по принадлежности к железам внутренней секреции - половые гормоны, тиреоидные гормоны и т.д. Классификация истинных гормонов: 1) пептидные и белковые гормоны - гипоталамуса и гипофиза (тиролиберин, соматолиберин, соматостатин, гормон роста, кортикотропин, тиреотропин) и поджелуд.жел (инсулин, глюкагон) 2) гормоны – производные аминокислот - мозгового вещества надпочечников (адреналин, норадреналин), и гормоны щитовидной железы (тироксин и его производные) IV уровень. Уровень целостного организма. Нервная и гуморальная регуляция соподчинены и сопряжены поведенческим реакциям. Классификация БАВ: 1.Неспецифические метаболиты (НМ) - продукты метаболизма, вырабатываемые любой клеткой в процессе жизнедеятельности и обладающие биологической активностью (СО2, молочная кислота). 2.Специфические метаболиты (СМ): а) тканевые гормоны (парагормоны); б) истинные гормоны. СМ-продукты жизнедеятельности, вырабатываемые определенными специализированными видами клеток, обладающие биологической активностью и специфичностью действия: а) тканевые гормоны - БАВ, вырабатывающиеся специализированными клетками, оказывают эффект в основном на месте выработки. Формируют т.н. APUD-систему Система тканевых гормонов: Система тканевых гормонов
Главные функции гормонов: 1. Метаболическая функция гормонов. Метаболическая функция гормонов – влияние на обмен веществ (анаболизм – соматотропный гормон, катаболизм – тироксин, адреналин). 2. Коррегирующая функция гормонов. Коррегирующая функция гормонов – регуляция текущей деятельности отдельных систем организма и их функций. 3. Кинетическая (триггерная) функция гормонов. Кинетическая (триггерная) функция гормонов – включение функций. Более частные, но очень важные функции гормонов: 1. Морфогенетическая функция гормонов. Морфогенетическая функция гормонов – влияние на морфогенез: рост, развитие (СТГ, тироксин, половые гормоны). 2. Адаптивная функция гормонов. Адаптивная функция гормонов – влияние на реакции приспособления к условиям существования во внешней среде (АКТГ, глюкокортикоиды). 3. Репродуктивная функция гормонов. Репродуктивная функция гормонов – влияние на функцию размножения (женский половой цикл, сперматогенез). На этом уровне регуляции парагормоны выполняют свою функцию при попадании в кровь, при этом они оказывают дистантное действие. 12. Гуморальная регуляция функций. Межсистемный уровень регуляции функций, роль истинных гормонов и парагормонов, классификация истинных гормонов. Понятие о нейрогуморальной регуляции. Различия между нервной и гуморальной регуляцией. Различия нервной и гуморальной регуляции 1. Действие гормонов, в связи с их транспортом кровью, более диффузно; нервное влияние более дифференцированно. 2. Гуморальное влияние распространяется медленнее (максимальная скорость кровотока - 0,2-0,5 м/сек). 3. Гуморальное влияние более длительное. Нейрогуморальная регуляция - совместное регулирующее, координирующее и интегрирующее влияние нервной системы и гуморальных факторов, содержащихся в крови, лимфе и тканевой жидкости биологически активных веществ — метаболитов, гормонов, медиаторов - на физиологические процессы в организме. Парагормоны - БАВ, вырабатывающиеся специализированными клетками, оказывают эффект в основном на месте выработки. Различия нервной и гуморальной регуляции. 1. Действие гормонов, в связи с их транспортом кровью, более диффузно; нервное влияние более дифференцированно. 2. Гуморальное влияние распространяется медленнее (максимальная скорость кровотока 0,2-0,5 м/сек). 3. Гуморальное влияние более длительное (часы, сутки). Истинные гормоны – БАВ, вырабатывающиеся в специализированных железах внутренней секреции, обладающие дистантным действием и высокой биологической активностью. Гормонпродуцирующие структуры. Все гормонпродуцирующие структуры делятся на: -гипофизарно-гипоталамическую систему; -периферические эндокринные железы; -специализированные эндокринные образования в неэндокринных органах; -диффузную эндокринную систему,состоящую из одиночных клеток с эндокринной функцией. Классификация гормонов. Гормоны делятся по: – принадлежности к железам внутренней секреции – половые гормоны, тиреоидные гормоны и т.д.; – химической природе на: - белковые (некоторые гормоны аденогипофиза); - пептидные (инсулин); - производные аминокислот (тироксин, адреналин, норадреналин); - стероидные производные холестерина (половые гормоны, гормоны коры надпочечников); - производные полиненасыщенных жирных кислот; – электрическому заряду на: -полярные, гидрофильные гормоны. К ним относятся белковые, пептидные гормоны и гормоны, производные аминокислот (кроме тиреоидных гормонов); -неполярные гидрофобные гормоны. К ним относятся стероиды (половые гормоны и гормоны коры надпочечников), производные полиненасыщенных жирных кислот (простагландины) и тиреоидные гормоны. 13. Гормональная регуляция. Понятие о гормонах, их классификация и свойства. APUD-система. Парагормоны. Типы функционального влияния гормонов. Механизмы действия гормонов, специфичность. Понятие об органах- и клетках-мишенях, клеточных рецепторах. Этапы образования и выделения белковых и пептидных гормонов. Формирование гормонального ответа на клеточном уровне. Механизм действия белковых гормонов. Комплекс гормон-рецептор. Системы вторичных посредников, их характеристика. Механизм действия стероидных гормонов, его отличительные особенности. Гормоны оказывают четыре типа воздействия: А) метаболическое – влияет на различные виды обмена веществ; Б) морфогенетическое действие – влияют на рост, развитие и дифференцировку тканей и органов, созревание организма; В) пусковое действие – активируют работу того или иного органа; Г) корригирующие действие – изменяют функции органов в соответствии с потребностями организма. Этапы образования и выделения белковых и пептидных гормонов: 1. биосинтез гормона; 2. секреция, т.е. выделение из эндокринной клетки; 3. транспорт кровью к периферическим тканям; 4. распознавание гормонального сигнала клетками-мишенями; 5. трансдукция (перевод) гормонального сигнала в биологический ответ; 6. гашение гормонального сигнала. Схема регуляции: уровень гормона - рецепторы ГТ - выработка рилизинг-факторов - кровь - аденоГФ - выработка тропных гормонов - кровь - железы - выработка гормонов - кровь - органы и ткани - рецепторы ГТ. Выделяют два механизма действия на клетки-мишени: 1. Водорастворимые гормоны взаимодействуют с рецепторами мембраны (катехоламины, пептидные и белковые гормоны), далее происходит активация внутримембранных аденилатциклаз и синтез цАМФ или цГМФ (универсальные вторичные посредники в процессе реализации эффекта гормона) из АТФ и ГТФ. Четыре системы 2-ных посредников 1. Система Аденилатциклаза – циклический Аденозинмонофосфат (цАМФ) активация протеинкиназы, сокращение мышечной кл 2. Система Гуанилатциклаза - циклический Гуанозинмонофосфат (цГМФ) расслабление мыш кл, фосфорилирование снижается Эффекты цАМФ и цГМФ - противоположны. 3. Система фосфолипаза С - инозитол-три-фосфат 4. Система Са++ - кальмодулин 2. Стероидные гормоны вследствие жирорастворимости проникают внутрь клетки, затем в ядро, где стимулируют или угнетают синтез белков и ферментов, влияя на генетический аппарат ядра (половые гормоны, гормоны коры надпочечников). 1.При контакте стероидного гормона с мембранным рецептором происходит: узнавание гормона; перенос ГРК в клетку. 2.В цитоплазме гормон связывается с особым (вторым) цитоплазматическим белком-рецептором с образованием ГРК. 3.Перенос ГРК в ядро. 4.Взаимодействие с ядерным (третьим) рецептором с образованием ГРК. 5.Связывание ГРК с хроматиновым акцептором и ДНК. 6.Активация транскрипции матричных РНК (мРНК), синтез транспортных и рибосомных РНК, трансляция мРНК, 7.Синтез регуляторных белков и ферментов в рибосомах. 8.Физиологические эффекты (наступают через несколько часов, т.к. требуют длительного присутствия ГРК в ядре). 14. Гипоталамо-гипофизарная система и ее роль в регуляции функций. Нейросекреция гипоталамуса. Понятие о рилизинг-факторах (либерины и статины). Пути взаимодействия гипоталамуса и гипофиза. Классификация гормонов гипофиза. Петли регуляции, их характеристика. ГТ является высшим центром, осуществляющим нейро-гуморальную регуляцию. Информация от рецепторов ГТ поступает в супраоптическое и паравентрикулярное ядра ГТ, нейроны которых обладают способностью к нейросекреции, что проявляется в изменении уровня выделения рилизинг-факторов – это группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза. Схема регуляции: уровень гормона - рецепторы ГТ - выработка рилизинг-факторов - кровь - аденоГФ - выработка тропных гормонов - кровь - железы - выработка гормонов - кровь - органы и ткани - рецепторы ГТ. С функциональной точки зрения рилизинг-факторы подразделяются на две группы: Либерины – это рилизинг-гормоны, способствующие усилению синтеза и секреции соответствующего гормона в эндокринных клетках передней доли гипофиза. Статины – это рилизинг-гормоны, подавляющие синтез и секрецию… В настоящее время известно 10 рилизинг-факторов (7 либеринов и 3 статина). Либерины: 1. Соматолиберин 2. Тиролиберин 3. Люлиберин 4. Фоллиберин 5. Кортиколиберин 6. Пролактолиберин 7. Меланолиберин Статины: 1. Соматостатин 2. Пролактостатин 3. Меланостатин Некоторые гормоны ГФ вырабатываются в ГТ и лишь накапливаются в ГФ (гормоны нейрогипофиза – Антидиуретический гормон (АДГ) и окситоцин). Регуляция выработки гормонов осуществляется по принципу отрицательной обратной связи: а. Ультракороткая петля регуляции: В самой железе есть рецепторы, которые измеряют концентрацию гормонов в крови. б. Короткая петля регуляции: Рецепторы ГФ реагируют на концентрацию тропных гормонов, а, следовательно, и периферических гормонов. в. Длинная петля регуляции: Уровень периферических гормонов воспринимается хеморецепторами ГТ - ГФ - ЖВС - гормон. Передняя, задняя и промежуточная доли гипофиза, их гормоны, физиологическое действие, механизмы действия на клетку. Гормоны аденогипофиза (передняя доля): Адренокортикотропный (АКТГ) - стимулирует работу коры надпочечников. Синтез глюкокортикоидов. по принципу обратной связи. Тиреотропный (ТТГ) - регулируется несколькими факторами: влиянием рилизинг-факторов гипоталамуса; отрицательной обратной связью; суточным ритмом – наибольшая концентрация ТТГ наблюдается ночью. Тиреотропин стимулирует работу щитовидной железы и синтез тироксина и трийодтиронина. Также под воздействием ТТГ активируется синтез белков, потребление йода, увеличивается размер тиреоидных клеток. Пролактин – гормон молочных желез. Он стимулирует их рост и развитие. Пролактин необходим для лактации – он вызывает образование молока после беременности. Дополнительно он отвечает за торможение овуляционного цикла. Это достигается благодаря подавлению секреции ФСГ. Регуляция: через ГТ и рефлекторно (пролактолиберин и пролактостатин). Фолликулостимулирующий (ФСГ) Выработка ФСГ регулируется гипоталамусом. действует на яичники и яички. У женщин ФСГ ускоряет развитие фолликулов и выработку эстрогенов. У мужчин влияет на клетки яичек – стимулирует сперматогенез. У женщин уровень ФСГ зависит от фазы менструального цикла. Лютеинизирующий (ЛГ) необходим для репродукции. В организме женщины под действием ЛГ происходит превращение остаточного фолликула в желтое тело. В дальнейшем желтое тело начинает выработку прогестерона – главного гормона беременности. У мужчин ЛГ влияет на клетки яичек, которые вырабатывают тестостерон. Тип секреции ФСГ и ЛГ у мужчин - тонический, у женщин - циклический. Гр. гландотропных гормонов. Регуляция:ГТ (фоллиберин, люлиберин)(ФСГ и ЛГ) Соматотропин (СТГ) гормон роста. относится кэффекторнымгормонам. Регуляция: ГТ (соматолиберин и соматостатин). Промежуточной доли: Меланоцитостимулирующий отвечает за выработку пигментов кожи, волос, сетчатки глаз. Регуляция: рилизинг-факторы ГТ (меланостатин и меланолиберин). Липотропин стимулирует липолиз. Основная функция липотропина заключается в образовании эндорфинов. Нейрогипофиз (задняя доля): В нейрогипофизе гормоны не вырабатываются, а только активируются и накапливаются вазопрессин (антидиуретический гормон гипофиза, АДГ) и окситоцин. Местом синтеза окситоцина и вазопрессина является гипоталамус. Щитовидная железа, ее гормоны, их классификация, характеристика, физиологические эффекты, механизмы действия на клетку. Регуляция гормональной функции щитовидной железы. Паращитовидные железы, их гормоны и физиологические эффекты. Вырабатываются два йодосодержащих гормона - трийодтиронин (Т3) и тироксин (Т4), один не содержащий йода - тиреокальцитонин. Функция щитовидной железы регулируется гипоталамо-гипофизарной системой по механизму обратной связи. В гипоталамусе секретируется гормон тиреолиберин, под влиянием которого стимулируется продукция аденогипофизом тиреотропина. Тиреотропин взаимодействует с рецепторами щ/ж-зы и стимулирует выработку тиреоидных гормонов. Секрецию тиреолиберина тормозит гормон гипоталамуса соматостатин, который угнетает также продукцию тиреотропина. Механизм обратной связи является основополагающим в деятельности эндокринных желез. Уровень тиреоидных гормонов в крови регулирует продукцию тиреолиберина и тиреотропина, который влияет на синтез тиреоидных гормонов. При снижении в крови уровня тиреоидных гормонов усиливается продукция тиреолиберина и тиреотропина, что повышает секрецию тиреоидных гормонов и поступление их в кровь. При повышении уровня тиреоидных гормонов в крови тормозится секреция тиреолиберина и тиреотропина и, соответственно, тиреоидных гормонов. Физиологические эффекты гормонов: 1. Участвуют в формировании адаптивных реакций. Повышение возбудимости ЦНС, активация психических процессов. 2. Повышают чувствительность тканей к катехоламинам; подавляет активность ферментов, разрушающих адреналин (моноаминоксидазы). 3. Активация симпатических эффектов (тахикардия, повышение сократимости миокарда, потливость). 4. Защитное действие от стрессорных повреждений миокарда и желудка (изъязвления). 5. Активируют теплообразование и теплоотдачу. 6. Усиление диуреза за счет увеличение почечного кровотока, клубочковой фильтрации и угнетения канальцевой реабсорбции. 7. Усиливают физиологическую регенерацию тканей. 8. Влияют на рост и развитие: Гипотиреоз. А) Недостаточность в детском возрасте ведет к нарушению умственного развития (кретинизм) Б) Микседема - снижение уровня Т3, Т4 в зрелом возрасте. Характеризуется снижением умственной и физической активности. В) Эндемический зоб - разрастание ткани щитовидной железы при недостаточном содержании в пище и воде йода по принципу отрицательной обратной связи: недостаток йода приводит к снижению концентрации гормонов, это сопровождается увеличением уровня ТТГ, который стимулирует ткань железы. Гипертиреоз (болезнь Грейвса, Базедова болезнь, диффузный токсический зоб)- в основе - образование Длительно действующего стимулятора щитовидной железы (LATS) - подобен ТТГ, следовательно, нет регуляции по принципу отрицательной обратной связи (аутоиммунное заболевание). Околощитовидные железы продуцируют паратгормон. При недостатке паратгормона понижается, а при избытке повышается содержание кальция в крови. Избыточная функция (гиперпаратиреоз) околощитовидных желез наблюдается довольно редко, например, при опухолях околощитовидных желез. При этом содержание кальция в крови увеличено, а количество неорганического фосфата уменьшено. Развивается остеопороз, т.е. разрушение костной ткани, мышечная слабость, вынуждающего больного постоянно лежать, боли в спине, ногах, руках. При недостаточности внутрисекреторной функции околощитовидных желез у человека (гипопаратиреоз), вследствие падения уровня кальция в крови резко повышается возбудимость центральной нервной системы, и возникают приступы судорог. Физиология надпочечников. Гормоны коры надпочечников, классификация, роль в регуляции функций организма, механизмы действия на клетку. Гормоны мозгового вещества надпочечников, их физиологическое значение. Регуляция гормональной активности надпочечников. По химическому составу активные вещества делятся на несколько групп: минералокортикоиды: альдостерон
глюкокортикостероиды: кортизол, кортизон
андрогены: тестостерон, эстроген, прогестерон
катехоламины: АД, Н-АД
пептиды: соматостатин.
Кора надпочечников имеет сложное строение, в ней выделяют три части: Клубочковая (поверхностный слой, самый тонкий). Пучковая (средняя). Сетчатая (примыкает к мозговому веществу). Важную роль в образовании глюкокортикоидов играет кортикотропин передней доли гипофиза. Это влияние осуществляется по принципу прямых и обратных связей: кортикотропин повышает продукцию глюкокортикоидов, а избыточное их содержание в крови приводит к торможению кортикотропина в гипофизе. В ядрах переднего отдела гипоталамуса синтезируется нейросекрет кортиколиберин, который стимулирует образование кортикотропина в передней доле гипофиза, а он, в свою очередь, стимулирует образование глюкокортикоида. Функциональное отношение «гипоталамус – передняя доля гипофиза – кора надпочечников» находится в единой гипоталамо-гипофизарно-надпочечниковой системе, которая играет ведущую роль в адаптационных реакциях организма. Эндокринная функция поджелудочной железы, ее гормоны, их характеристика и роль в регуляции обмена веществ, механизмы действия на клетку. Клетки поджелудочной железы, синтезирующие гормональноактивные вещества, называются инсулоцитами. Они расположены в железе скоплениями – островками Лангерганса. По строению различают несколько типов инсулоцитов: альфа, бета, дельта, РР и эпсилон. Поджелудочная железа вырабатывает пять гормонов. Два из них существенно влияют на обмен веществ. Это инсулин и глюкагон. Другие гомоны имеют меньшее значение для регуляции метаболизма, либо секретируются поджелудочной железой в малых количествах. Инсулин Анаболический гормон, основной функцией которого является транспорт сахара в клетки организма. Он снижает уровень глюкозы. Глюкагон Принимает важнейшее участие в углеводном обмене. Основная функция этого гормона поджелудочной железы – стимуляция гликогенолиза (процесс расщепления гликогена, в процессе которого в кровь выделяется глюкоза). Женские половые железы. Женские половые гормоны, их характеристика, физиологическое значение, механизмы действия на клетку. Эндокринная функция плаценты. В плаценте из эстрона образуется эстриол, кроме того плацента синтезирует прогестерон, которые выполняют присущие им функции (см. половые гормоны) а так же хорионический гонадотропин, который участвует в регуляции дифференцировки и развития плода, также влияет на организм матери, вызывая: задержку воды и солей, усиление секреции вазопрессина, активацию механизмов иммунитета. Мужские половые железы. Мужские половые гормоны, их характеристика, физиологическое значение, механизмы действия на клетку. |