Рефлексы. Нарушения чувствительности. Топическая диагностика. Классификация
 Скачать 1.44 Mb. Скачать 1.44 Mb.
|

|
Теория воротного контроля боли(R.Melzak, P.Wall,1965): суть – центральный контроль за афферентным входом. Функцию входных ворот выполняет желатиновая субстанция задних рогов, модулирующая синаптическую передачу нервных импульсов с периферических волокон к центральным клеткам. Substantia gelatinosa – функциональная единица, простирающая вдоль спинного мозга; клетки связаны друг с другом короткими волокнами и длинными волокнами- тракт Лиссауэра, но не выходят за пределы студенистого вещества. Активность клеток студенистого вещества модулирует мембранный потенциал окончаний афферентных волокон и обусловливает возбудительный эффект приходящих импульсов. Это фоновая активность – проводится тонкими миелиновыми и немиелинизированными волокнами. Толстые волокна более эффективны в активации Т-клеток Имеется временная и пространственная суммация или интеграция Т-клетками приходящих импульсов. Контроль над сенсорным входом осуществляют задние столбы и дорсолатеральный путь. А-волокна входят в спинной мозг и посылают короткие ветви к желатиновой субстанции и длинные центральные ветви к ядрам дорсальных столбов, которые образуют медиальную петлю. 1-я система – система задних столбов, несет различение двух точек , различие силы, пространственной локализации, тактильных порогов и вибрации. 2-я система – дорсо-латеральный путь- начинается в задних рогах и проецируется после переключения в латеральном шейном ядре в ствол мозга и таламус. Очень быстрое проведение, по скорости опережает медиальную петлю. Система имеет четко ограниченные рецептивные поля и очень быстрое проведение. Обе эти системы выполняют функцию центрального контролбного триггера; они несут точную информацию о природе и локализации стимула;осуществляют подготовку восприятия корковых нейронов для последующих афферентных волн; воздействие при помощи центрального контроля эфферентных волокон на систему контроля ворот; избирательно активируют мозговые процессы. Любое повреждение, ухудшающее нормирование нисходящего потока импульсов к системе воротного контроля боли, вскрывает ворота. При недостаточности тормозных механизмов Т-клеток происходит их растормаживание и активация различными стимулами с периферии и из других источников, что ведет к интенсивной восходящей импульсации. Крыжановский Г.Н.(1976) – теория “генераторных механизмов “; процесс генерирования собственной активности структурами мозга; на уровне межнейрональных и системных отношений. В системе болевой чувствительности начинают действовать гиперактивные нейроны – ГПУВ. Поэтому можно использовать для ингибирования ГПУВ тормозные медиаторы – ГАМК, глицин; а также блокаторы кальциевых каналов –верапамил, магний. Патологическая алгическая система – патофизиологическая основа болевого синдрома (Г.Н.Крыжановский). Уровни патологической алгической системы (Г.Н.Крыжановский): 1)Периферический отдел – сенситизированные ноцицепторы, очаги эктопического возбуждения (поврежденные и регенерирующие невормы, демиелинизированные участки нерва), группа гипреактивных спинальных ганглиев; 2)Спинальный уровень – ГПУВ в афферентных ноцицептивных реле – в дорсальных рогах и ядрах спинального тракта тройничного нерва; 3) Супраспинальный уровень – ядра ретикулярной формации ствола, ядра таламуса, сенсомторная и орбитофронтальная кора, эмоциогенные структуры. В ПАС- взаимопддержание и взаимопотенциирование прямыми и обратными связями (порочные круги). Боль проводится по Аb-волокнам(25м/с) и С-волокнам(1м/с) к нейронам 1 и 5 слоев заднего рога. Висцеральные болевые импульсы поступают к клеткам 7 и 8 слоев. Спино-таламический путь делится на неоспиноталамический( несет информацию о боли сразу в таламус) и палеоспиноталамический( может опосредовать вегетативные и аффективные реакции на боль). Проекции палеоспиноталамического пути: а) на n. gigantocellularis – имеею широкие билатеральные рецептивные поля, отвечают потенциально болевые стимулы и также на зрительные и слуховые стимулы; то есть эти ядра участвуют в общем повышении внимания и ориентации на болевые стимулы; б) в среднем мозга на n. cuneiformis – от 1 и 5 слоев спинного мозга. Система восприятия боли на таламическом уровне организованно в основном контралатерально. В таламусе заканчиваются аксоны 1,4,5,7,8 слоев. Медиальный компонент спиноталамического тракта дает проекцию в парафасцикулярное и центральное таламические ядра. Каудальная часть таламуса- латеральный компонент спиноталамического тракта. Нейрофизиологические феномены боли при стимуляции С-волокон:
Нейрохимические механизмы боли:
Боль – это своеобразное психофизиологическое состояние человека, возникающее в результате воздействия сверхсильных или разрушительных раздражителей, вызывающих органические или функциональные нарушения в организме. Это не только ощущение, но также интегративная функция организма, которая мобилизует самые разнообразные функциональные системы для защиты организма от воздействия вредного фактора. Биологический смысл боли по И. П.. Павлову, состоит в отбрасывании и выбрасывании всего, что мешает (угрожает) жизненному процессу, что нарушило бы равновесие организма со средой. Боль – сигнал опасности. Согласно наиболее распространенному мнению, один из компонентов синдрома боли – ощущение – возникает при возбуждении неинкапсулированных нервных окончаний, которые по сути своей являются хеморецепторами. При действии повреждающих агентов кинины и некоторые ионы (кальций, в норме находящийся внутри клеток) попадают в межклеточные пространства и раздражают нервные окончания, обладающие низким порогом возбудимости. Далее ноцицептивное раздражение, проводится преимущественно по безмиелиновым и по тонким миелиновым волокнам. Известно, что выделяют два вида болевых ощущений: быструю локальную или эпикритическую боль и медленную, задержанную, протопатическую (эволюционо более древнюю). Проводятся они по разным путям. Процесс передачи и обработки возбуждений, формирующих боль, обеспечивается структурами, расположенными на разных уровнях нервной системы. Следует отметить, что болевые импульсы передаются и по путям вегетативной нервной системы, в частности, через пограничный ствол. Поступление болевой импульсации в ЦНС и ее переработка происходят по механизмам с обратной связью. Первый уровень локализуется в желатинозной субстанции задних рогов спинного мозга, где регулируется приток импульсов к клеткам задних рогов. Мелзак и Уолл (1965) на основании своих исследований выдвинули теорию «контроля афферентного потока на выходе». По спиноталамическому тракту возбуждение достигает специфических ядер зрительного бугра (особенно его вентролатеральной группы). Ноцицептивные сигналы протопатической чувствительности проводятся по экстралемнисковым системам – спиноретикулярному, спинотектальному и спинобульбарному путям. На стволовом уровне большое значение для проведения и интеграции ноцицептивных сигналов имеют ядра шва (средний мозг), а далее импульсы идут в вентролатеральные ядра зрительного бугра. Ведущая роль таламуса в формировании болевых ощущений подтверждается современными данными. В нем ноцицептивное раздражение, пришедшее с периферии, приобретает характер первичного ощущения. Далее, в болевой интеграции принимает участие лимбическая система мозга, имеющая отношение к памяти и эмоциям. Боль как интегративная функция включает и такие компоненты, как сознание, ощущение, память, мотивация, вегетативные, соматические и поведенческие реакции. В оформлении болевого импульса играет роль циркуляции импульсов по таламокортикальным круговым орбитам. В процессе реверсации происходит считывание пришедшего возбуждения с имеющейся генетической информацией. Процесс осознания боли как ощущения, локализации ее по отношению к определенной области тела и весь комплекс поведения, связанный с болью, совершаются при обязательном участии коры больших полушарий и, прежде всего, соматосенсорной зоны. Активация, возникающая при ноцицептивном раздражении, формируется с участием ретикулярной формации ствола, особенно мезэнцефальной области. По данным Melzak (1965), одним из модуляторов болевого раздражения являются задние канатики спинного мозга, в которых расположены пути глубокой чувствительности. Именно эти пути содержат быстро проводящие миелинизированные волокна, позволяющие головному мозгу быстро получать информацию о характере и локализации внешних воздействий. Увеличение активности задних канатиков тормозит передачу боли из клеток заднего рога. Именно поэтому при избирательном повреждении первых возникает резко выраженные боли. 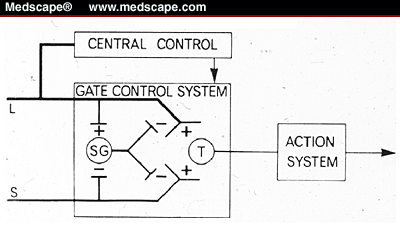 The Gate Control Model. Large-diameter afferent fibers (L) stimulate the substantia gelatinosa (SG) and the transmission cells (T) in lamina 4. The SG cells reduce the membrane potential of afferent terminals, thus producing presynaptic inhibition. Small-diameter afferent fibers (S) also stimulate the transmission cells but inhibit SG cells and thus turn off the existing presynaptic inhibition. Published with permission from: Melzack R, Wall P. Pain mechanisms: a new theory. Science. 1965;150:975. Терапия боли. ДВИГАТЕЛЬНАЯ СИСТЕМА. Движения непроизвольные (рефлекторные) и произвольные (с участием коры головного мозга). Двигательная система осуществляет регуляцию произвольной и непроизвольной моторики. Двигательная система включает в себя афферентную часть – кинестетическая основа, которая обеспечивается задними(постцентральными) отделами коркового ядра двигательного анализатора и перерабатывает экстероцептивные сигналы в кинестетические, а также эфферентную часть, представленную передними отделами коркового ядра двигательного анализатора(осуществляет серию последовательных иннерваций, протекающих во времени; “кинетическая мелодия“). Первый двигательный нейрон локализуется в коре передней центральной извилины, в задних отделах верхней и средней лобных извилин, парацентральной дольке. При поражении эфферентной части анализатора выпадают тонкие движения (только пирамидный тракт поражается по Скупченко), снижение силы движений происходит, развиваются параличи. При поражении афферентного отдела доминантного полушария страдает нужный набор кинестетических импульсов, осуществляющих движение; “афферентный парез “ – теряется нужный мышечный адресат, диффузное сокращение агонистов и антагонистов, нет достаточной дифференцировки движений; явления статической атаксии, то есть кинестетическая апраксия, которая максимально проявляется в контралатеральной руке, но может выявляться и в одноименной руке. Проекционная двигательная зона состоит:1) первичная проекционная двигательная зона – начало пирамидного пути, прецентральная извилина (корковое поле№4); 2) вторичная проекционная двигательная зона – происходит интеграция эфферентных двигательных импульсов; локализуется в средней и верхней лобных извилинах (задние отделы)- поля №6 и8; 3) третичная проекционная двигательная зона – осуществляется сложный синтез двигательных импульсов, необходимых для выполнения четких, плавных и высокоорганизованных двигательных актов; 10 коркове поле; передние отделы нижней и средней лобных извилин. Иннервация мускулатуры перекрестная; проекция клеток – обратное расположение частей тела – нижние конечности в верхнем слое извилины, верхние конечности – нижний слой извилины. Неспособность осуществлять целесообразные движения при отсутствии парезов, атаксии или нарушении тонуса – апраксия (Липманн,1906). Второй двигательный нейрон – периферический: а) двигательные ядра черепно-мозговых нервов; б) клетки передних рогов спинного мозга. Клетки передних рогов спинного мозга находятся под контролем следующих структур:
Двигательные клетки передних рогов спинного мозга – конечный общий путь(Sherington,1904). Функция мышц и их иннервация
Функция мышц туловища и их иннервация
Бернштейн И.А. (1990) указывает, что двигательная система представляет собой кинематическую цепь с различными степенями свободы,состоящую из: 1) пассивной части – жесткого скелета и суставов; 2) активной части – поперечно-полосатой мускулатуры со всем ее оснащением. Как нам кажется, в это определение следует добавлять и управленческую часть (головной мозг) и проводниковую часть (проводящие пути головного и спинного мозга, нервы). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||