биология. EKZAMEN_BIO (Восстановлен). Краткий обзор 1) единство химического состава, 2) обмен веществ, 3) самовоспроизведение (репродукция), 4) наследственность
 Скачать 1.8 Mb. Скачать 1.8 Mb.
|

|
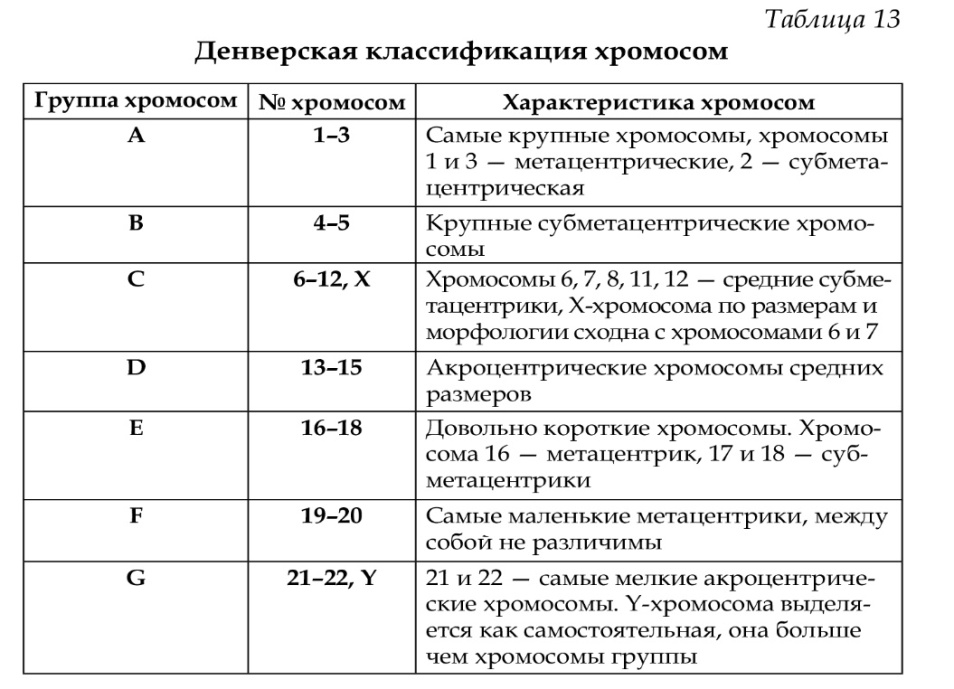
Первый уровень укладки ДНК в хроматине обеспечивается нуклеосомами. Они представляют собой округлые частицы диаметром 15 нм, которые связаны между собой участками ДНК длиной около 20 нм. Отдельная нуклеосома состоит их белковой сердцевины, на которую накручена молекула ДНК. Белковая сердцевина нуклеосомы, или кор, имеет форму диска диаметром 11 нм и толщиной 6 нм. Она содержит по две молекулы гистонов H2A, H2B, H3 и H4. Если развернуть сердцевину, то можно обнаружить, что молекулы гистонов соединены в последовательности H2A, H2B, H4, H3, H3, H4, H2B, H2A. При сворачивании сердцевины молекулы гистонов располагаются как бы в два этажа, наподобие винтовой лестницы. Молекула ДНК в виде левозакрученной суперспирали совершает 1,75 оборота вокруг сердцевины. При этом в непосредственный контакт с гистонами вступает 146 пар нуклеотидов. Длина линкерной ДНК, соединяющей соседние нуклеосомы, колеблется в пределах 40-70 пар нуклеотидов в зависимости от типа клетки. Таким образом, на одну нуклеосому приходится в среднем около 200 пар нуклеотидов ДНК. Нуклеосомы укорачивают молекулу ДНК примерно в 7 раз. Они обнаружены во всех эукариотических клетках и даже у ДНК-содержащих вирусов. Однако это не означает, что вся ДНК клеточного ядра связана с нуклеосомами, и некодирующие повторы могут иметь другую укладку. Второй уровень укладки ДНК обеспечивается взаимодействием линкерной ДНК с гистоном H1. Молекула гистона H1 своим глобулярным доменом связывается с двумя витками ДНК на нуклеосоме. Одновременно C-концевой фибриллярный домен гистона H1 вступает в контакт с линкерной ДНК. В результате соседние нуклеосомы приближаются друг к другу, формируя группы из 6-8 частиц – нуклеомеры (супербусины)диаметром 25-30 нм. Последовательность аминокислот в C-домене гистона H1 гомологична первичной структуре некоторых регуляторов транскрипции. В связи с этим предполагается, что гистон H1 конкурирует с факторами транскрипции за связывание с линкерной ДНК, контролируя тем самым активность генов. Третий уровень укладки ДНКпредставлен хроматиновыми фибрилламидиаметром 30 нм, которые хорошо видны в электронном микроскопе в интерфазных ядрах и митотических хромосомах. Они имеют суперспиральную структуру и содержат максимально сближенные между собой нуклеомеры. В формировании фибрилл диаметром 30 нм принимают участие гистон H1, а также негистоновые белки с HMG-доменом. За конденсацию хроматина в фибриллу диаметром 30 нм отвечает, прежде всего, C-концевой домен гистона H1. При этом первостепенное значение приобретает уже не взаимодействие гистона с ДНК, а взаимные связи гистоновых молекул между собой. Образование нуклеомеров и хроматиновых нитей диаметром 30 нм вызывает дальнейшую компактизацию ДНК в 40-50 раз. Четвертый уровень укладки ДНКобеспечивается взаимодействием фибрилл диаметром 30 нм с ядерным матриксом. При этом формируются петлевые домены, содержащие в среднем 90 тысяч пар нуклеотидов. В расправленном состоянии их длина может достигать 20 мкм. Концы таких петель прикреплены к ядерному матриксу в особых точках, обозначаемых как MARs (matrix attachment regions) или SARs (scaffold attachment regions). Эти точки содержат молекулы свободной ДНК (“форум-ДНК”) длиной 50-150 пар нуклеотидов, которые устойчивы к действию нуклеаз и выделяются из ядра независимо от высокомолекулярной ДНК. Точки MARs содержат также топоизомеразу II, участвующую в формировании изгибов ДНК. Со стороны ядерного матрикса прикрепление хроматиновых нитей обеспечивается ламином A. Петлевые домены являются типовой структурно-функциональной единицей хроматина. Домен во многом автономен, независимо реплицируется и транскрибируется. В своем составе домены имеют кластеры генов, которые связаны функционально. Петлевые домены обеспечивают компактизацию молекулы ДНК в 700 раз. Пятый уровень укладки ДНКсвязан с формированием групп из 18-20 петлевых доменов, прикрепленных в виде розетки к общему центру из белков ядерного матрикса. Розетки из петлевых доменов находятся в хроматине в компактном состоянии, образуя округлые гранулы диаметром около 150 нм – хромомеры. При активации локализованных в хромомере генов его величина может возрастать до 300 нм и более. Шестой уровень укладки ДНКопределяется формированием хромонемы – фибриллярной структуры диаметром 200-300 нм, состоящей из плотно упакованных хромомеров.В хроматине интерфазного ядра хромонемы обычно не выявляются. Они становятся видимыми только при конденсации хроматина в профазе митоза, а также в ранней телофазе при деконденсации хромосом. Хромомерный и хромонемный уровни укладки позволяют укоротить длину молекулы ДНК в 10 000 раз. Седьмой уровень укладки ДНКсостоит в образовании хроматид (однохроматидных хромосом) из хромонем. Толщина хроматиды составляет в среднем 700-800 нм. Если учесть, что толщина хромонемы обычно равна 100-200 нм, то коэффициент упаковки для хромосомного уровня составляет не более 10. Способ укладки хромонемы в хромосоме изучен недостаточно. У одних видов хромонема имеет вид спирали, у других в одной хромосоме могут обнаруживаться две и более параллельные друг другу хромонемы.Хромосомный уровень укладки ДНК в большей степени, чем другие уровни, отражает видовые особенности организации генома эукариот. Каждый вид, как известно, имеет характерный для него набор хромосом. Однако хромосомы можно наблюдать в микроскоп только при делении клеток митозом или мейозом. В ядрах неделящихся клеток хромосомы находятся в деконденсированном состоянии, образуя хроматин. Тем не менее, даже в интерфазном ядре хромосомы сохраняют свою индивидуальность, занимая в нем определенные хромосомные территории. Согласно К. Раблю (1885) хромосомы прикреплены к нуклеолемме теломерными концами, тогда как центромерные участки располагаются ближе к центру ядра. Новейшие исследования интерфазного ядра с помощью конфокальной микроскопии подтверждают эту концепцию, добавляя к ней ряд существенных деталей. В частности, гомологичные хромосомы локализованы на противоположных сторонах ядра. При активации генов и сопутствующей этому деконденсации хромосомы удлиняются, смещаясь к центру ядра. Вот почему в животных и некоторых растительных клетках гетерохроматин концентрируется преимущественно по периферии, тогда как эухроматин занимает центральную область ядра. ВОПРОС № 105 Принцип, лежащий в основе Международной Денверской классификации хромосом человека Принципы: 1) различия в длине хромосом 2) расположение центромеры Хромосомы располагаются в порядке уменьшения их длины. Все пары хромосом распределены на 7 групп и пронумерованы арабскими цифрами: группа А(1-3) – большие метацентрические, В(4-5) – большие субметацентрические, С(6-12) – средние субметацентрические и Х-хромосома, D(13-15) – средние акроцентрические, Е(16-18) – небольшие мета- и субцентрические, F(19- 20) – самые короткие метацентрические, G(21-22) – самые мелкие акроцентрические, Y-хромосома. Центромерный индекс отражает длину короткого плеча к длине всей хромосомы. ОСНОВНАЯ ЧАСТЬ Основы существующей унифицированной классификации хромосом были заложены в 1960 году в Денвере. В основу классификации положены различия в длине хромосом и расположении центромеры. На основании различий в длине выделены 23 пары хромосом, при этом парам, имеющим наибольшую длину, дан наименьший номер (самыми длинными являются хромосомы 1- и 2-й пары). Выделяют группы метацентрических, субметацентрических и акроцентрических хромосом. Отнесение хромосом к тому или иному типу производится на основе расчета центромерного индекса - отношения длины короткого плеча к длине всей хромосомы. В группе метацентрических хромосом короткое и длинное плечи приблизительно равны, и центромерный индекс приближается к 0,5. В субметацентрических хромосомах центромерный индекс снижен и составляет от 0,25 до 0,35, в акроцентрических хромосомах он часто не превышает 0,2. На основании комбинации этих двух основных признаков хромосомы сгруппированы в 7 групп, обозначаемых буквами английского алфавита (от А до G).
Вопрос 106. Международная Парижская классификация хромосом человека. Краткий обзор: Латинскими буквами р и q обозначаются соответственно короткое и длинное плечо хромосомы. От центромеры к теломере по имеющимся отчетливым морфологическим указателям (маркерам) в каждом плече выделяют районы, обозначаемые арабскими цифрами. В пределах районов идентифицируют сегменты — регулярные участки, отличающиеся по интенсификации окраски. Они также обозначаются арабскими цифрами. Основная часть: Парижская классификация. В1971 году в Париже на IV международном конгрессе по генетике человека была согласована единая система идентификации хромосом человека, учитывавшая дифференцировку хромосом по длине.Каждая хромосома набора человека при дифференциальной окраске характеризуется уникальным для нее сочетанием темно окрашенных сегментов или полос , чередующихся с неокрашенными участками или светлыми сегментами. Именно такое специфическое для данной хромосомы сочетание сегментов позволяет четко ее идентифицировать и отличить от других хромосом набора. В пределах короткого (р) и длинного (q) плеча каждой хромосомы выделяют ряд четко идентифицируемых областей или регионов, которые нумеруются арабскими цифрами начиная от центромеры к теломерному участку или терминальному концу хромосомы. Каждая область хромосомы включает определенное число сегментов, нумерация которых (второй арабской цифрой) также идет в направлении от центромерного к теломерному участку. Таким образом, обозначение хромосомного сегмента 2q34 означает хромосому №2, длинное плечо, 3 регион и 4 сегмент. Сама центромера обозначается сочетанием цифр 1 и 0, т.е. часть центромеры в пределах короткого плеча обозначается как- р10, а часть, включающая длинное плечо -q10 Открытие в середине 70-х годов того факта, что профазные и про-метафазные хромосомы позволяют достичь большего числа сегментов, чем метафазные хромосомы, и, следовательно, повысить разрешающие возможности цитогенетического исследования, привело к разработке методов получения хромосом высокого разрешения и потребовало дополнения цитогенетической номенклатуры новыми принципами анализа таких хромосом. В1980 году по этому поводу в Париже было достигнуто международное соглашение, которое было опубликовано в 1981 году под названием "Международная система цитогенетической номенклатуры хромосом человека - сегментация хромосом высокого разрешения" или ISCN (1981). Так, если сегмент в пределах какой-либо хромосомы подразделяется на отдельные субсегменты, то после номера сегмента ставится точка, после которой указывается номер субсегмента. Например, если оригинальный сегмент 1 р31 подразделяется на 3 разных субсегмента, то они обозначаются как 1р31.1, 1р31.2и 1р31.3, причем субсегмент 1р31.1 является проксимальным, а 1 р31.3 - дистальным по отношению к центромере. Дополнительное деление субсегментов на другие сегменты, например субсегмента 1 р31.1, соответственно обозначается как 1p31.11,1р31.12 ит.д. Вопрос №107 Назовите основные виды хромосомных аберраций. Краткий обзор: Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — изменения структуры хромосом. Классифицируют делеции (удаление участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую). Хромосомные перестройки носят, как правило, патологический характер и нередко приводят к гибели организма. Показано значение хромосомных перестроек в видообразовании и эволюции Основная часть: Делеции (от лат. deletio — уничтожение) — хромосомные перестройки, при которых происходит потеря участка хромосомы. Делеция может быть следствием разрыва хромосомы или результатом неравного кроссинговера. По положению утерянного участка хромосомы делеции классифицируют на внутренние (интерстициальные) и концевые (терминальные). Инверсия - изменение структуры хромосомы, вызванное поворотом на 180° одного из внутренних её участков. Дупликация (лат. duplicatio — удвоение) — мутация, нарушающая структуру хромосом, представляет собой удвоение участка хромосомы, содержащего гены. Может произойти в результате ошибки при гомологичной рекомбинации, Ретротранспозиции, или из-за дубликации всех хромосомы.[1] Вторая копия гена часто не подвергается давлению селекции — так, мутация одной из копий гена не несет вреда организму. Следовательно, копии накапливают мутации быстрее, чем гены, существующие в одном экземпляре. Транслока́ция — тип хромосомных мутаций, при которых происходит перенос участка хромосомы на негомологичную хромосому. Отдельно выделяют реципрокные транслокации, при которых происходит взаимный обмен участками между негомологичными хромосомами, и Робертсоновские транслокации, или центрические слияния, при которых происходит слияние акроцентрических хромосом с полной или частичной утратой материала коротких плеч. Особый вид реципрокных транслокаций представляют собой так называемые робертсоновские транслокации. В этом случае разрывы в двух акроцентрических хромосомах локализуются в области центромер или в непосредственной близости от них. Длинные плечи хромосом сливаются, а короткие теряются. Поскольку короткие плечи акроцентрических хромосом содержат гены рРНК, то их потеря никак не проявляется, так как множественные копии этих генов содержатся также в других акроцентрических хромосомах. Поэтому робертсоновская транслокация функционально является сбалансированной. Реципрокные транслокации являются сбалансированной хромосомной перестройкой, при их формировании не происходит потери генетического материала. Вопрос 108. Генные мутации. Механизмы их возникновения. Краткий ответ: Генные точечные мутации характеризуются искажением порядка следование нуклеотидов в составе гена-мишени, что может выразиться прекращением трансляции белковой молекулы, синтезом мутантного белка с нарушенной функцией или токсическими свойствами, недостаточным или избыточным синтезом белка. По последствиям генных мутаций их классифицируют на нейтральные и динамические, а также на миссенс- и нонсенс-мутации. (см. ниже) Полный ответ: По характеру изменений в составе гена различают следующие типы мутаций: Делеции — утрата сегмента ДНК размером от одного нуклеотида до гена. Дупликации — удвоение или повторное дублирование участка ДНК от одного нуклеотида до целых генов. Инверсии — встраивание от двух до нескольких нуклеотидов на прежнее место ДНК после поворота на 180 Инсерции — вставка фрагментов ДНК размером от одного нуклеотида до целого гена. Трансверсии — замена пуринового (А,Г) основания на пиримидиновое (Ц,У,Т) или наоборот в одном из кодонов. Транзиции — в нуклеотидном остатке замена одного пуринового основания на другое пуриновое или одного пиримидинового на другое в структуре кодона. Нейтральная мутации (молчащая мутация) — мутация не имеет фенотипического выражения (например, в результате вырожденности генетического кода). Миссенс-мутация — замена нуклеотида в кодирующей части гена, что приводит к замене аминокислоты в полипептиде. Нонсенс-мутация — замена нуклеотида в кодирующей части гена — приводит к образованию стоп-кодона и прекращению трансляции. Динамические мутации — мутации, обусловленные увеличением числа тринуклеотидных повторов в функционально значимых частях гена. Такие мутации могут привести к торможению или блокаде транскрипции, приобретению белковыми молекулами свойств, нарушающих их нормальный метаболизм. Вопрос 109. Виды генных мутаций Краткий ответ: Генные мутации – изменение строения одного гена. Это изменение в последовательности нуклеотидов: выпадение, вставка, замена и т.п. Например, замена А на Т. Причины – нарушения при удвоении (репликации) ДНК. Примеры: серповидноклеточная анемия, фенилкетонурия. Генные, или точечные, мутации бывают в основном 3 видов: замены, при которых одно азотистое основание в ДНК замещается на другое; вставки, обеспечивающие внедрение в молекулу ДНК одного или нескольких дополнительных нуклеотидов; делеции (или выпадения) одного или нескольких нуклеотидов, при которых происходит укорочение молекулы ДНК. Полный ответ: Генные мутации – изменение строения одного гена. Это изменение в последовательности нуклеотидов: выпадение, вставка, замена и т.п. Например, замена А на Т. Причины – нарушения при удвоении (репликации) ДНК. Примеры: серповидноклеточная анемия, фенилкетонурия. По характеру изменений в составе гена различают следующие типы мутаций: Делеции — утрата сегмента ДНК размером от одного нуклеотида до гена. Дупликации — удвоение или повторное дублирование сегмента ДНК от одного нуклеотида до целых генов. Инверсии — поворот на 180° сегмента ДНК размером от двух нуклеотидов до фрагмента, включающего несколько генов. Инсерции — вставка фрагментов ДНК размером от одного нуклеотида до целого гена. Трансверсии — замена пуринового основания на пиримидиновое или наоборот в одном из кодонов. Транзиции — замена одного пуринового основания на другое пуриновое или одного пиримидинового на другое в структуре кодона. По последствиям генных мутаций их классифицируют на нейтральные, регу-ляторные и динамические, а также на миссенс- и нонсенс-мутации. Нейтральная мутации (молчащая мутация) — мутация не имеет фенотипи-ческого выражения (например, в результате вырожденности генетического кода). Миссенс-мутация — замена нуклеотида в кодирующей части гена — приводит к замене аминокислоты в полипептиде. Нонсенс-мутация — замена нуклеотида в кодирующей части гена — приводит к образованию кодона-терминатора (стоп-кодона) и прекращению трансляции. Регуляторная мутация — мутация в 5'- или З'-нетранслируемых областях гена, такая мутация нарушает экспрессию гена. Динамические мутации — мутации, обусловленные увеличением числа три-нуклеотидных повторов в функционально значимых частях гена. Такие мутации могут привести к торможению или блокаде транскрипции, приобретению белковыми молекулами свойств, нарушающих их нормальный метаболизм. |