Курс лекций Естествознание
 Скачать 3.99 Mb. Скачать 3.99 Mb.
|

|
1. Свойства и функции корня Корень – один из основных вегетативных органов растения, в типичном случае выполняющий функцию водного и минерального питания. Корень – осевой орган, которому характерны следующие свойства: – цилиндрическая форма; – радиальная симметрия; – неограниченный верхушечный рост за счет деятельности апикальной меристемы; – положительный геотропизм(рост в направлении вектора силы тяжести); – отсутствие листьев и цветов; – наличие в редких случаях эндогенных придаточных почек, из которых развиваются придаточные побеги; – интенсивное ветвление и огромная внешняя поверхность, позволяющая более полно освоить почвенное пространство. Рис. 5.1. Строение молодого растения конских бобов. Корни, как и все другие органы, поглощают кислород и выделяют углекислый газ. В растении корень выполняет следующие функции: – питательная – поглощение из субстрата воды и растворенных в ней минеральных веществ; – механическая – закрепление растения в субстрате (почве и др.); – запасающая – в корнях многих растений откладываются запасные питательные вещества; – синтез различных веществ (аминокислоты, алкалоиды, гормоны (гиббереллины, цитокинины) и др.), поступающих затем в другие органы растения; – взаимодействует с корнями других растений (посредством выделения веществ – аллелопатия), вступает в мутуалистический симбиоз с грибами (микориза) и клубеньковыми бактериями (клубеньки на корнях бобовых и др.); – является органом вегетативного размножения (благодаря придаточным почкам из которых вырастают побеги – корневые отпрыски, например, у розы, терна, сирени, осины, осота, малины и др.); – иногда выполняет роль дыхательного органа (пневматохоры мангровых зарослей). 2. Виды корней и корневых систем По происхождению выделяют три типа корней (рис. 5.1.; 5.2.): – главный корень – первый корень семенного растения, развивающийся из зародышевого корешка. В почве занимает вертикальное положение и, как правило, толще остальных; успоровых растений (плаунов, хвощей и папоротников) его нет; – придаточный корень – корень, берущий начало от других органов растения: стеблей (кукуруза, ива), листьев (бегония), корневищ (ландыш, осоки), клубней (картофель), луковиц (лук); – боковой корень – ответвление главного или придаточного корней. Отличается слабо выраженным геотропизмом – растет горизонтально или косо вниз. Совокупность всех корней одного растения, независимо от их происхождения и структуры называется корневой системой. Чаще всего выделяют три типа корневых систем: – система главного корня, или стержневая – корневая система с хорошо выраженным главным корнем, а также с его ответвлениями – боковыми корнями. Характерна для большинства голосеменных (ель, сосна) и древесных двудольных растений (береза, тополь, осина), многих трав (одуванчик, морковь). В некоторых случаях главный корень не разветвляется или ветвится мало – простой корень (морковь, ряска); – система придаточных корней, или мочковатая – корневая система, представленная в основном придаточными корнями, а также боковыми. Такая система встречается у покрытосеменных однодольных растений (злаки, осоки, ситники) и некоторых двудольных (лютики);  – система главного и придаточных корней, или смешанная – корневая система, представленная как хорошо развитым главным, так и придаточными корнями, а также боковыми. Характерна некоторым двудольным растениям (подсолнечник, фасоль, земляника и др.). В зависимости от того субстрата, в котором расположены корни и откуда они извлекают воду и питательные вещества, различают четыре экологических типа корней: – подземные – целиком или хотя бы частично расположены в грунте (у 70%высших растений); – водяные – всегда расположены в толще воды и никогда не достигают дна водоема (ряска, многокоренник, водокрас); – воздушные – находятся в воздухе, поглощают атмосферную влагу. Например, у растений-эпифитов – обитателей влажных тропиков (орхидеи, монстера); – гаустории – корни-присоски паразитических растений (повилика, омела), эпифитных лиан (плющ). Некоторые растения корней вообще не имеют: мхи (вместо корней развиты ризоиды – экзогенные выросты покровной ткани; у сфагновых мхов и их нет), некоторые водные растения (папоротник сальвиния, вольфия бескорневая (самое маленькое цветковое растение на нашей планете), пузырчатка). 3. Внешнее строение корня Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон корня (рис. 5.3.): –  зона деления – апикальная меристема корня (от 1 до 5 мм), в результате интенсивного деления ее клеток формируются все прочие зоны и ткани корня; от повреждений защищена корневым чехликом – постоянно обновляющимся образованием на верхушке растущего корня – представлен особой паренхимной тканью, которая защищает апикальную меристему от трения о почвенные частицы и способствует продвижению корня благодаря выделению слизи; водные растения, как правило, корневого чехлика не имеют (вместо него формируется чехликоподобное образование – корневой кармашек); зона деления – апикальная меристема корня (от 1 до 5 мм), в результате интенсивного деления ее клеток формируются все прочие зоны и ткани корня; от повреждений защищена корневым чехликом – постоянно обновляющимся образованием на верхушке растущего корня – представлен особой паренхимной тканью, которая защищает апикальную меристему от трения о почвенные частицы и способствует продвижению корня благодаря выделению слизи; водные растения, как правило, корневого чехлика не имеют (вместо него формируется чехликоподобное образование – корневой кармашек);– Рис. 5.3. Общий вид (А) и продольный раз- рез (схема Б) молодого корешка: I – корневой чехлик, прикрывающий зону деления; II – зона роста; III – зона всасывания; IV – зона проведения; 1 – корневой чехлик, 2 – апекс корня, 3 – калиптроген, 4 – корневые волоски, 5, 6 – боковые корни. зона роста, или растяжения – зона роста клеток (несколько мм), растягиваясь в продольном направлении, проталкивает корневое окончание вглубь почвы; клетки зоны характеризуются высоким тургором; в этой зоне начинается дифференциация клеток; – зона всасывания (от нескольких мм до нескольких см) ответственна за поглощение воды и минеральных веществ при помощи корневых волосков – выростов клеток эпиблемы (ризодермы) продолжительность жизни корневых волосков – 10–20 дней; У одного растения ржи примерно 14 млрд. корневых волосков суммарной длиной более 10 000 км. – зона проведения обладает хорошо развитой проводящей тканью, передает почвенные растворы в вышележащие отделы растения; корневые волоски вместе с клетками ризодермы погибают, и формируется покровная ткань, защищающая живые ткани корня; здесь появляются боковые корни (отсюда и второе название зоны – зона ветвления); составляет основную массу корня. Резких границ между зонами корня нет. Место перехода корня в стебель называется корневой шейкой. 4. Внутреннее (первичное и вторичное) строение корня Снаружи молодые корневые окончания покрыты эпиблемой (ризодермой) – однослойной первичной покровной тканью корня (образуется из дерматогена – наружный слой верхушечной меристемы кончика корня) (рис. 5.4.). В зоне поглощения клетки эпиблемы образуют корневые волоски, а в зоне проведения они довольно быстро слущиваются. Проникновение воды и солей в корневые волоски и далее в клетки коры и центрального цилиндра происходит путем осмоса, диффузии и активного переноса. П 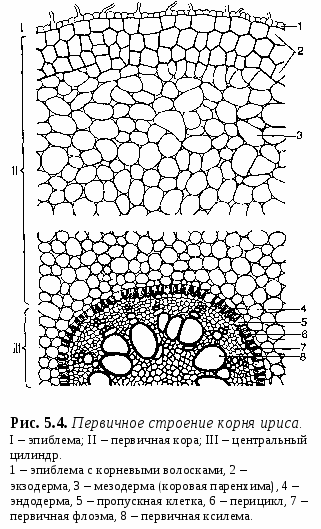 од эпиблемой располагается первичная кора (дифференцируется из периблемы – периферийный отдел верхушечной меристемы, лежащий глубже дерматогена). од эпиблемой располагается первичная кора (дифференцируется из периблемы – периферийный отдел верхушечной меристемы, лежащий глубже дерматогена).Наружные клетки первичной коры, лежащие непосредственно под эпиблемой, называются экзодермой. Она бывает однослойной и многослойной (2 – 3 слоя). В зоне проведения после слущивания эпиблемы экзодерма оказывается снаружи, может опробковевать и выполняет функцию защитной покровной ткани. О  сновная масса первичной коры – мезодерма –образована паренхимными клетками: многослойная, рыхлая, с межклетниками. Через нее проходит радиальный (ближний) транспорт воды с минеральными веществами от эпиблемы к центральному цилиндру; здесь осуществляется активный синтез метаболитов и откладываются запасные питательные вещества. сновная масса первичной коры – мезодерма –образована паренхимными клетками: многослойная, рыхлая, с межклетниками. Через нее проходит радиальный (ближний) транспорт воды с минеральными веществами от эпиблемы к центральному цилиндру; здесь осуществляется активный синтез метаболитов и откладываются запасные питательные вещества.Самым внутренним слоем коры является эндодерма, которая выполняет роль барьера – контролирует передвижение веществ из коры в осевой цилиндр и обратно (рис. 5.5.). На ранних этапах развития эндодерма состоит из живых, тонкостенных клеток, расположенных плотно, без межклетников. Позднее ее клетки приобретают некоторые характерные структурные особенности: на радиальных стенках клеток эндодермы появляются особые утолщения (в результате суберинизации и одревеснения) – пояски Каспари, с помощью которых перекрывается передвижение растворов вдоль клеточных стенок (апопластный путь). В результате этого все вещества в центральный цилиндр и из него могут проникать только через живые протопласты клеток эндодермы (симпластный путь) и под их контролем (избирательная проницаемость, защита от проникновения болезнетворных микроорганизмов). У однодольных в клетках эндодермы могут происходить дальнейшие изменения: на внутренней поверхности первичных оболочек клеток откладывается суберин и далее вторичная целлюлозная оболочка, которая со временем одревесневает, таким образом перекрывается и второй путь передвижения веществ – симпластный (через цитоплазму клеток). На некоторых клетках этого не происходит и они остаются только с поясками Каспари, это так называемые пропускные клетки – у однодольных только через них осуществляется физиологическая связь между первичной корой и осевым цилиндром. Пропускных клеток в эндодерме мало, а ее диаметр намного меньше, чем внешний диаметр корня в области эпиблемы, поэтому вокруг эндодермы образуется повышенное в сравнении с другими тканями количество воды. В результате этого вода проникает через пропускные клетки в сосуды ксилемы с определенным давлением, которое получило название корневого давления (рис. 5.6.). 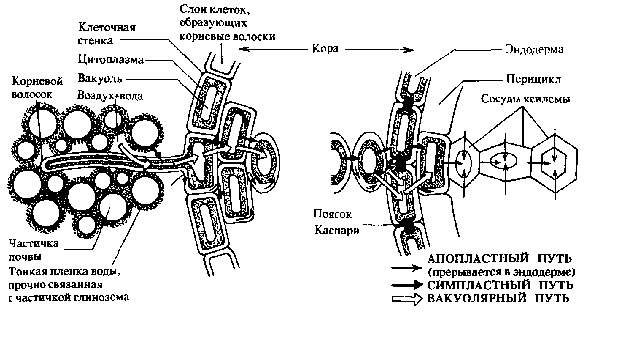 Рис. 5.6. Схема передвижения воды в корне. Центральный (осевой) цилиндр (формируется из плеромы – внутренней части апикальной меристемы) начинает дифференцироваться в зоне роста, вплотную к зоне деления. Наружный его слой – перицикл представляет собой однослойную образовательную ткань, состоящую из живых паренхимных клеток. В перицикле закладываются боковые корни, а также у некоторых растений возникают зачатки придаточных почек. У двудольных растений он участвует во вторичном утолщении корня, отчасти образуя камбий и феллоген. Центральную часть цилиндра занимает сосудистоволокнистый пучок, состоящий из первичной ксилемы и первичной флоэмы (проводящие ткани образуются из прокамбия, который закладывается под перициклом). Элементы флоэмы и ксилемы закладываются по кругу, чередуясь друг с другом, и развиваются центростремительно (по направлению к центру корня), однако ксилема растет быстрее и занимает центр корня – сформировавшаяся структура проводящей ткани получила название радиального проводящего пучка. Сердцевина не типична для корня, но иногда (например, у кукурузы) заметна в центре в виде небольшого участка механической ткани или тонкостенных клеток, возникающих из прокамбия. Такое первичное строение корня сохраняется до конца жизни у хвощей, плаунов, папоротников и однодольных, а у остальных – только в зоне всасывания. Для голосеменных и двудольных растений характерны вторичные изменения в строении корня, обеспечивающие его рост в толщину. Вторичные изменения происходят за счет вторичных меристем – камбия и феллогена (рис. 5.7.). В 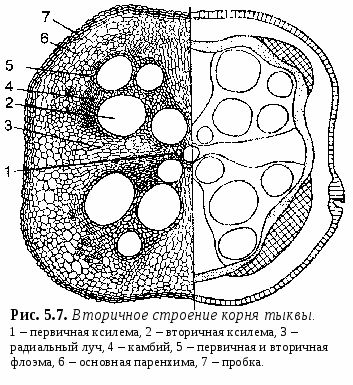 начале камбий появляется из клеток паренхимы центрального цилиндра (прокамбия) между сосудами первичной ксилемы и первичной флоэмы (с внутренней стороны). Участки камбия постепенно разрастаются, огибают флоэму и соприкасаются с перициклом. Клетки камбия прокамбиального происхождения делятся и дифференцируются: к центру откладывается вторичная ксилема, к наружи – вторичная флоэма. В местах, где сосуды первичной ксилемы соприкасаются с перициклом, клетки последнего также превращаются в камбий (перициклического происхождения), но он дифференцируется только в паренхиму, которая образует радиальные лучи напротив первичной ксилемы. Образуется камбиальное кольцо. начале камбий появляется из клеток паренхимы центрального цилиндра (прокамбия) между сосудами первичной ксилемы и первичной флоэмы (с внутренней стороны). Участки камбия постепенно разрастаются, огибают флоэму и соприкасаются с перициклом. Клетки камбия прокамбиального происхождения делятся и дифференцируются: к центру откладывается вторичная ксилема, к наружи – вторичная флоэма. В местах, где сосуды первичной ксилемы соприкасаются с перициклом, клетки последнего также превращаются в камбий (перициклического происхождения), но он дифференцируется только в паренхиму, которая образует радиальные лучи напротив первичной ксилемы. Образуется камбиальное кольцо.Вследствие быстрого нарастания изнутри вторичных тканей, обусловливающего сильное утолщение корня, первичная кора нередко разрывается. Перицикл также становится меристематически активным: он делится и в центробежном направлении откладывает пробковый камбий – феллоген. Феллоген делится и кнаружи откладывает пробку (феллему) – вторичную покровную ткань корня. Пробка изолирует первичную кору от проводящих тканей, она теряет связь с живыми клетками центрального цилиндра, отмирает и слущивается. Внутрь феллоген откладывает феллодерму, которая затем превращается в паренхимную ткань и образует вместе с перициклом вторичную кору. Снаружи корни двудольных растений, имеющие вторичное строение, покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев. Многолетние корни древесных растений в результате длительной камбиальной активности нередко сильно утолщаются. Вторичная ксилема у таких корней сливается в сплошной цилиндр, окруженный снаружи кольцом камбия и сплошным кольцом вторичной флоэмы. Таким образом, этапы перехода корня от первичного строения ко вторичному следующие: 1. Появление камбия между лучами ксилемы и флоэмы и образование камбиального кольца; 2. Образование феллогена перициклом, возникновение вторичной коры; 3. Сбрасывание первичной коры; 4. Смена радиального расположения тканей ксилемы и флоэмы коллатеральным. 5. Основные видоизменения корня (метаморфозы) Большинство растений имеют корни типичного строения. Но у многих видов в процессе эволюции корни приспособились к выполнению особых функций, в связи с чем, строение их изменилось. Такие изменения называются метаморфозами. З 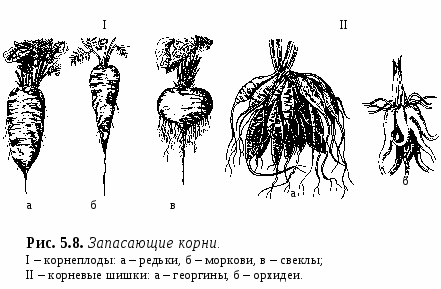 апасающие корни. У многолетних растений запасные питательные вещества могут откладываться и в корнях. Если функция запаса становится основной, то такие корни называются запасающими. По происхождению и структуре различают два типа запасающих корней: корнеплоды и корневые шишки (рис. 5.8.). апасающие корни. У многолетних растений запасные питательные вещества могут откладываться и в корнях. Если функция запаса становится основной, то такие корни называются запасающими. По происхождению и структуре различают два типа запасающих корней: корнеплоды и корневые шишки (рис. 5.8.).Корнеплоды образуются за счет разрастания главного корня. В образовании корнеплода принимает участие нижняя часть стебля, причем у свеклы, репы, редиса она составляет большую часть корнеплода, а собственно корень – только нижняя его часть, на котором развиваются боковые корни. Запасные продукты у корнеплодов (крахмал, инулин, различные сахара) могут откладываться в паренхиме вторичной коры (морковь, петрушка) или в паренхиме древесины (редька, репа, редис). Изредка запасные вещества откладываются в паренхиме, образованной деятельностью нескольких добавочных колец камбия (свекла) – пример третичного строения (установлено, что образование добавочных камбиальных колец стимулируется деятельностью листьев – их число примерно равно числу листьев, деленному на два). Корневые шишки (корневые клубни)возникают при разрастании боковых корней (у георгина, чистяка, ятрышника, батата). Образовывают придаточные почки и служат не только для перезимовки, но и для вегетативного размножения. М  икориза. Микориза представляет собой мутуалистический симбиоз корней многих растений с гифами грибов (некоторые зигомикоты и аскомикоты, но главным образом базидиальные грибы) (рис. 5.9.). Грибной компонент облегчает корням получение воды и минеральных веществ из почвы, а также, видимо, передает им некоторые органические вещества. Гриб в свою очередь получает от растения углеводы и другие питательные вещества. икориза. Микориза представляет собой мутуалистический симбиоз корней многих растений с гифами грибов (некоторые зигомикоты и аскомикоты, но главным образом базидиальные грибы) (рис. 5.9.). Грибной компонент облегчает корням получение воды и минеральных веществ из почвы, а также, видимо, передает им некоторые органические вещества. Гриб в свою очередь получает от растения углеводы и другие питательные вещества. Р  азличают эктотрофную микоризу, когда гифы гриба охватывают корень только снаружи, иногда проникая в межклетники коровой паренхимы (сосна, береза, дуб, ива и др.), и эндотрофную микоризу, когда грибной чехол вокруг корня не образуется, а гифы проникают глубоко в корень и внедряются в клетки коровой паренхимы (яблоня, груша, земляника, злаки, орхидные и др. – характерна для большинства покрытосеменных). В а б в Рис. 5.10. Втягивающие корни. а – крокуса, б – кислицы, в – лилии; 1 – втягивающие корни. а б в тягивающие или контрактильные корни. Характерны для многих корневищных и луковичных растений, обитающих чаще всего в экстремальных условиях (регионы с сухим жарким климатом или суровой зимой), а также характерны для многих сельскохозяйственных растений (клевер, гречиха, люцерна, морковь, свекла). Укорачиваясь у основания, такие корни (более длинные, чем обычные) способны втягивать луковицы или корневища в почву на оптимальную для их сохранения глубину (рис. 5.10.). К 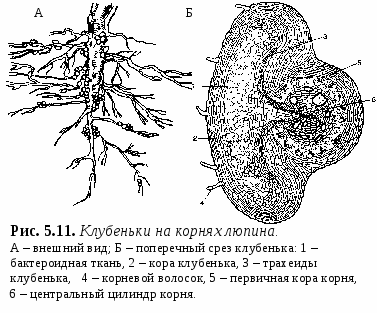 лубеньки. Клубеньки представляют собой разрастание паренхимной ткани корня (у многих бобовых), вызванное некоторыми видами бактерий из рода Rhizobium (рис. 5.11). Эти микроорганизмы способны фиксировать атмосферный молекулярный азот, переводя его в связанное легкоусвояемое растением состояние. С другой стороны, бактерии используют вещества, находящиеся в корнях растения, а также стимулируют разрастание корней. Благодаря данному симбиозу происходит обогащение почвы азотом. лубеньки. Клубеньки представляют собой разрастание паренхимной ткани корня (у многих бобовых), вызванное некоторыми видами бактерий из рода Rhizobium (рис. 5.11). Эти микроорганизмы способны фиксировать атмосферный молекулярный азот, переводя его в связанное легкоусвояемое растением состояние. С другой стороны, бактерии используют вещества, находящиеся в корнях растения, а также стимулируют разрастание корней. Благодаря данному симбиозу происходит обогащение почвы азотом.На корнях ольхи, лоха, облепихи также образуются клубеньки, однако они возникают в результате симбиоза с актиномикотами, которые также способны фиксировать атмосферный азот. Корни-гаустории, или присоски. Такие корни характерны для паразитических растений (омела, заразиха, повилика и др.), эпифитных лиан (плющ). У  омелы, например, паразитирующей на стволах и ветвях различных древесных растений, образуются длинные цилиндрические или слегка сплюснутые корни, растущие в коре растения-хозяина (рис. 5.12.). От них в проводящую ткань отходят присоски, которые поглощают его питательные вещества (воду и минеральные вещества). Из придаточных почек на этих корнях образуются побеги, пробивающиеся через кору наружу. В |