Курс лекций по зоологии. Курс лекций РостовнаДону 2010 южный федеральный университет
 Скачать 7.18 Mb. Скачать 7.18 Mb.
|

|
Ктенофорная гипотеза предполагает, что появление бескишечных турбеллярий объясняется дальнейшей редукцией пищеварительной системы, в ходе которой произошла редукция глотки, а кишечник, имевший полость и внеклеточное пищеварение, сменился совокупностью аморфно расположенных клеток, осуществляющих внутриклеточное пищеварение. Если эволюционные изменения, постулируемые предыдущими положениями ктенофорной гипотезы, возможны (другой вопрос - имели ли они место в ходе реальной эволюции), то последнее предположение, безусловно, неверно. Редукция пищеварительной системы в эволюции многоклеточных животных встречается неоднократно, но она связана с переходом к паразитизму (у погонофор - к питанию за счет веществ, синтезированных симбиотическими бактериями). Неизвестно случаев редукции пищеварительной системы у свободноживущих, активно подвижных животных, хищников. У турбеллярий не имелось экологических причин для редукции кишечника. Кроме того, там, где кишечник редуцирован, питание не совершается за счет фагоцитоза, способом приема пищи для таких животных становится абсорбция питательных субстанций. Можно также видеть, что эта гипотеза не объясняет необособленный характер эпителия и заднее положение рта. Планулоидная гипотеза впервые была высказана Л.Граффом (серия работ 1888 – 1904 гг.). Эту гипотезу разделяли многие исследователи. Существенные дополнения в планулоидную гипотезу внесли В.Н.Беклемишев (1964) и А.В.Иванов (1968, 1973 и другие работы). Нельзя не упомянуть также и И.И.Мечникова (1887, 1886), который отмечал, что у бескишечных турбеллярий имеется аморфный фагоцитобласт, и этим они близки к гипотетической фагоцителле. Л.Графф в своих работах показал глубокую примитивность бескишечных турбеллярий и тем самым определил их место как предков всех остальных турбеллярий. Признаки примитивного строения бескишечных турбеллярий были перечислены ранее. Признавая главным из них отсутствие полости кишечника, Л.Графф писал: "Поскольку все турбеллярии проходят стадию стеррогаструлы с паренхимой, содержащей еще необособленные элементы энто- и мезодермы, нет сомнения в том, что предкам турбеллярий должна быть приписана ацелия (отсутствие кишечника), как примитивный признак, и что современные бескишечные турбеллярии ближе всего стоят к этим предкам". Итак, планулоидная гипотеза признает наиболее примитивными (плезиоморфными) бескишечных турбеллярий. В счвою очередь все примитивные особенности организации бескишечных турбеллярий объясняются унаследованностью от крайне примитивно организованных предков. Активное движение турбеллярий, хищничество, увеличение размеров - все это обусловило прогрессивную эволюцию турбеллярий, прогрессивное развитие группы. Различные способы решения одних и тех же задач, асинхронность развития разных систем в разных эволюционных стволах привели к формированию нескольких различающихся эволюционно продвинутых апоморфных групп турбеллярий. Что касается вопроса о том, от каких предков взяли начало сами бескишечные турбеллярии, то в более ранних вариантах этой гипотезы в качестве таковых рассматривались планулы (правильнее - паренхимулы) кишечнополостных. Для этих личиночных стадий характерно наличие переднезадней полярности, ресничного эпителия, аморфного состояния внутреннего пласта клеток. Их аборальный полюс находится на переднем конце тела, оральный - на заднем. После окончания периода свободного существования планулы садятся на субстрат передним концом (аборальным полюсом), а на заднем конце - на оральном полюсе - прорывается рот. Предполагается, что процесс эволюционного становления турбеллярий был запущен двумя характерными изменениями планул: 1) приобретение ими способности к размножению на стадии личинки (такое явление нередко встречается у животных и называется неотенией); 2) переходом планул к ползанию, к жизни на поверхности субстрата. В этом последнем случае планулы ложились на субстрат на одну из антимер (или, выражаясь еще проще, - на бок, эта часть тела становилась брюшной стороной животного). Можно выразиться и иначе: при таком способе оседания сначала никакой кардинальной перестройки организации не происходило, передний конец продолжал оставаться передним, задний - задним, главная ось и при плавании, и при ползании одновременно являлась и переднезадней осью. В результате этого перехода стал складываться турбеллярный план строения (прежде всего - формирование билатеральной симметрии и уплощенной формы тела), при этом бескишечные турбеллярии как бы остановились на более раннем этапе, тогда как остальные турбеллярии прошли по этому пути значительно дальше. Вопрос о предках бескишечных турбеллярий может быть решен и иначе. Возможно, что непосредственными их предками были фагоцителлобразные организмы - фагоцителлы первого уровня, то есть такие, которые уже сформировали на оральном полюсе рот, но их фагоцитобласт еще не был эпителизован, и пищеварение происходило только внутриклеточно. Через такую стадию прошли в своей эволюции кишечнополостные, поэтому паренхимула кишечнополостных рекапитулирует их основные черты. Сам же процесс перехода к организации турбеллярий от фагоцителл мыслится шедшим по описанной выше схеме. Теперь следует остановиться на еще одном кардинальном отличии ктенофорной и планулоидной гипотез. И в том и в другом случае толчком к формированию нового турбеллярного плана строения явилось оседание предка на субстрат, переход от плавания к ползанию. С точки зрения ктенофорной гипотезы оседание совершалось на оральный полюс. В результате изменения формы тела (сплющивание) главная, или аборально-оральная ось укорачивается, формируются две новые поверхности: аборальная, обращенная вверх, она становится физиологически спинной стороной, и оральная, обращенная к субстрату, она становится физиологически брюшной стороной. Продольная (переднее-задняя) ось ползающего гребневика является новацией, возникшей при переходе к ползанию. Если животное лежит на ровной поверхности грунта, то его главная ось направлена перпендикулярно к субстрату. С точки зрения планулоидной гипотезы оседание на субстрат проходило на одну из антимер, на бок. В этом случае укорачивания главной оси не происходит, она, как и при плавании, одновременно является продольной (переднее-задней) осью и расположена параллельно поверхности субстрата. Получается, что положение главной оси должно быть веским доводом в пользу одной из гипотез. Однако, напомним, что главная ось является зоологической абстракцией и, конечно, непосредственному обнаружению недоступна. Тем не менее у радиальносимметричных пелагических гребневиков мы обнаруживаем наличие меридиональных нервных сгущений, вытянутых от орального полюса к аборальному. Эти сгущения нервных клеток радиальносимметрично расположены вокруг главной оси. Меридиональные стволы ортогона турбеллярий вытянуты от переднего конца тела к заднему и у более примитивных форм радиальносимметрично расположены вокруг переднезадней оси. Это сопоставление позволяет убедиться в том, что главная ось гребневиков гомологична переднезадней оси турбеллярий, что эта последняя является главной осью животных. Таким образом, анализ положения главной оси у турбеллярий и гребневиков заставляет нас сделать выбор в пользу планулоидной гипотезы, признавая, однако, за предковую форму не собственно планул, а фагоцителл. Таким образом современная точка зрения на происхождение турбеллярий в очень сжатом виде выглядит следующим образом. Предками турбеллярий были фагоцителлобразные плавающие существа, у которых был сформирован рот, но фагоцитобласт был еще неэпителизован, а пища переваривалась внутриклеточно. При оседании таких фагоцителл на субстрат на антимеру возникала организация, характерная для бескишечных турбеллярий. В дальнейшем имела место прогрессивная эволюция первичных плоских червей, главнейшие направления которой заключались в формировании полости кишечника и перемещения рта вперед, а также прогрессивного развития нервной, мышечной и выделительной систем. План организации турбеллярий оказался весьма продуктивным в эволюционном отношении, от них взяли начало все остальные плоские черви, что доказывается наличием у всекх плоских червей единого плана строения. От них же взяли начало немательминты и немертины. Суть второй группы гипотез сводится к тому, что турбеллярии произошли от предка, имевшего целом и кровеносную систему в результате вторичного упрощения. Сторонники этих гипотез полагают, что главным фактором, обусловившим подобную трансформацию, могло стать уменьшение размеров тела, которое само по себе могло привести к утрате целомической полости и утрате транспортных систем (кроме диффузии). Турбеллярии, как предполагается этими гипотезами, взяли начало не от взрослых целомических животных, а от их личинок, то есть в становлении турбеллярий имела место неотения. Личинки целомических животных обладают малыми размерами, у них, как правило, сильно развиты ресничные образования, а целом и транспортные системы до поры до времени не развиты. Предполагается, что именно эти особенности унаследовали турбеллярии. Тем не менее, вдумчивый слушатель уже отметил, что в этой гипотезе не объясняются причины заднего положения рта и отсутствия пищеварительной полости. В заключение считаю целесообразным привести цитату из руководства Э.Рупперта и соавторов "неопределенное систематическое положение турбеллярий (как и всего таксона Плоские черви) пока не удается прояснить, используя средства молекулярной систематики и компьютерной нумерической таксономии". Справедливости ради следует отметить, что этот же источник несколькими страницами раньше пишет "последние данные молекулярной систематики тоже свидетельствуют не в пользу традиционных взглядов" (под традиционными взгядами понимается морфологическая точка зрения, в частности планулоидная теория). Так что консенсус относительно происхождения турбеллярий остается не достигнутым, а студентам предлагается самостоятельно оценить доводы представленных гипотез. Задание для самостоятельной работ: представьте себе, что свободно живущее животное начинает осваивать паразитический образ жизни. Определите, к каким новым факторам новой внешней среды этот организм должен приспособиться при переходе к паразитированию в кишечнике. Данное задание для самостоятельной работы является подготовкой студента к изучению адаптаций паразитов к их среде обитания. NB! При выполнении задания не следует забегать вперед и описывать адаптации паразитов. ТЕМА 3.27 КЛАСС ТРЕМАТОДЫ ИЛИ СОСАЛЬЩИКИ (TREMATODA)
Как уже указывалось, огромное большинство представителей типа Плоские черви являются паразитами. Трематоды на разных фазах жизненного цикла паразитируют у разных хозяев: двуполое поколение трематод паразитирует у позвоночных, партеногенетические поколения - преимущественно приурочено к брюхоногим моллюскам. Типовой размер двуполого поколения трематод оценивается примерно в 10-12 миллиметров, есть виды меньших размеров - до 2 миллиметров, есть виды, достигающие в длину 5-7 сантиметров (печеночные двуустки). Остальные стадии жизненного цикла трематод характеризуются микроскопическими размерами. Общая численность трематод в настоящее время оценивается в руководстве Э.Рупперта с соавторами (2008) приблизительно в 11000 видов, а в руководстве Вестхайде и Ригера (2008) - в 7200 видов. Трематоды как Плоские черви. Трематоды, а также другие паразитические представители плоских червей могут быть охарактеризованы как плоские черви, адаптированные к паразитизму. Главнейшие признаки типа Плоских червей у трематод выражены вполне отчетливо (лучше всего - у марит) и проявляются в общем плане строения тела: в форме тела, паренхиматозности, наличии замкнутой пищеварительной системы, ортогональной нервной системы, протонефридиальной выделительной системы, гермафодитизма (только одно семейство трематод - Кровяные двуустки - характеризуется раздельнополостью, причем этот признак явно является вторичным). Для всех паразитических плоских червей характерно также формирование сложного яйца (смотри раздел 3.20). Общая схема жизненного цикла трематод. Трематоды от остальных плоских червей, в том числе и паразитических, отличаются сложным жизненным циклом, протекающим по типу гетерогонии. В жизненном цикле этих червей правильно сменяют друг друга гермафродитное поколение, размножающееся двуполым способом, приуроченное к позвоночным хозяевам - марита и два поколения, размножающихся партеногенетически и приуроченных к моллюскам - спороциста и редия (в жизненных циклах некоторых трематод место редии занимает дочерняя спороциста). В жизненном цикле трематод также присутствуют две личинки: мирацидий, который является личинкой первого партеногенетического поколения, и церкария, которая является личинкой гермафродитного поколения. В огромном большинстве жизненных циклов имеется также стадия инцистированной церкарии. Гермафродитное (в случае кровяных двуусток - раздельнополое) поколение трематод обитает в позвоночных животных (окончательный хозяин). В результате двуполого размножения формируются "яйца", которые выходят во внешнюю среду. Выход "яиц" чаще всего совершается естественным путем, вместе с экскрементами, но если мариты локализованы не в кишечном тракте, то могут использоваться особые приемы, обеспечивающие выход "яиц". Иногда развитие зародыша начинается еще в матке материнского организма, так что к моменту выхода "яиц" из организма окончательного хозяина в них уже содержится сформированная личинка, но значительно чаще эти процессы происходят во внешней среде, но под оболочкой яйца, есть и промежуточные варианты, при которых развитие начинается в матке матери, а завершаентся во внешней среде. Как бы то ни было, в конечном итоге внутри яйца формируется стадия, которая называется мирацидий. Мирацидий выходит из яйца в воду. Мирацидий совершает активные поисковые движения и находит (если повезет) подходящего хозяина, в которого внедряется (активный путь заражения). Этого хозяина мы называем первым промежуточным хозяином. Таким хозяином практически у всех трематод являются моллюски, чаще всего брюхоногие. Мирацидий, внедрившись в хозяина, превращается в спороцисту. Превращение сопровождается морфологической перестройкой организации, таким образом, мирацидия мы должны оценить как личинку спорооцисты. Еще в организме мирацидия находилась группа рыхло расположенных клеток - зародышевых шаров, эти клетки в организме спороцисты начинают дробиться без оплодотворения. За дроблением следует дифференцировка клеток, и формируется следующая стадия - редия. Процесс формирования редий происходит без оплодотворения и поэтому оценивается как партеногенез, а спороциста может быть оценена как партеногенетическая самка. Редия также способная к партеногенезу, в результате которого формируется следующая стадия жизненного цикла - церкария. Церкарии покадают организм первого промежуточного хозяина. Церкарии активно плавают, а затем проникают в тело какого-либо вида животного и инцистируются. Эта последняя категория хозяев получает название второго промежуточного хозяина. Заражение окончательного хозяина осуществляется путем проглатывания инцистированнной лицинки. Для разных трематод характерен разный набор вторых промежуточных хозяев, но практически всегда эти хозяева являются типичной пищей окончательных хозяев. После проглатывания церкарий в организме окончательного хозяина превращается в мариту, то есть церкарий должен быть оценен как личинка мариты. Для большей наглядности приведем общую схему жизненного цикла трематод.  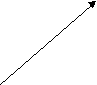 марита маритадвуполая стадия, 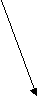 в окончательном хозяине=позвоночном в окончательном хозяине=позвоночномцеркария личинка, во внешней среде, позднее инцистируется 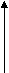 "яйцо" "яйцо"во внешней среде 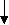 редия мирацидий партеногенетическая стадия, личинка, во внешней среде  в первом промежуточном в первом промежуточном  хозяине=моллюске хозяине=моллюске спороциста партеногенетическая стадия, в первом промежуточном хозяине=моллюске Жизненные чиклы конкретных видов трематод могут отличаться от этой общей схемы. Главнейшими из вариантов являются следующие: 1. Яйцо из организма окончательного хозяина должно попасть не в воду, а в наземную среду, первым промежуточным хозяином являются наземные виды моллюсков, в этом случае вылупления мирацидиев не происходит, мирацидий как свободная стадий отсутствует (пример - ланцетовидная двуустка). 2. И у тех видов трематод, развитие которых связано с водой, в некоторых случаях мирацидий не вылупляется, заражение первого промежуточного хозяина происходит путем проглатывания яйца, то есть пассивным для трематоды путем (пример - кошачья двуустка, китайская двуустка). 3. В результате партеногенеза спороцисты формируется не редия, а стадия, которая называется дочерней спороцистой (пример - кровяная двуустка). 4. Церкарии могут не покидать первого промежуточного хозяина, заражение окончательного хозяина происходит при поедании первого промежуточного хозяина (пример Leucochloridium paradoxum). У этого вида трематод церкарии скапливаются в щупальцах янтарки (наземная улитка). Заражение окончательного хозяина - певчих птиц - может произойти при проглатывании моллюска целиком, или в том случае, если птица оторвет и проглотит только щупальце янтарки. Щупальца зараженного моллюска приобретают яркую окраску, что представляет пример демаскировки хозяина, выгодной для паразита. 5. Церкарии не подвергаются инцистированию, они отыскивают окончательного хозяина и внедряются в его теле перкутанно (пример - кровяная двуустка). 6. Церкарии могут инцистироваться на поверхности животных или растений, а не в организме второго промежуточного хозяина. Заражение окончательного хозяина проихсодит путем проглатывания инцистированного церкария вместе с пищевым объектом или за счет случайного проглатывания (пример - печеночная двуустка). Характеристика стадий жизненного цикла трематод. Особенности организации марит трематод. Одной из особенностей трематод является наличие у них двух присосок, из которых первая помещается терминально на переднем конце тела, в глубине этой ротовой присоски находится рот. Вторая присоска называется брюшной, она размещена, понятно, на брюшной стороне тела, но положение ее у разных видов варьирует. Чаще всего она размещается на границе первой четверти или трети тела, но может быть размещена в задней половине тела и даже, например, у рода Diplodiscus (паразитирует в задней кишке лягушек) - терминально на заднем конце тела. Ранее полагали, что ротовое отверстие имеется и в брюшной присоске, с чем и связано название "двуустки", в учебно-научной литературе все чаще вместо термина "двуустки" пользуются термином "сосальщики". Для трематод (а также и для всех остальных паразитических плоских червей) характерно наличие покрова особого типа. В русскоязычной литературе обычно покров такого типа называют тегументом, в аглоязычной - неодермисом. На определенной стадии развития у паразитических плоских червей присутствует типичный ресничный эпителий. В какой-то момент жизненного цикла этот покров сбрасывается и замещается выростами необластов (смотри раздел 3.20). При этом тела клеток, которые называются цитоны и в которых лежит ядро, остаются в паренхиме и непосредственно граничат с клетками паренхимы и мускульными клетками. Отростки необластов пронизывают базальную пластинку и разрастаются по ее внешней поверхности, причем отростки соседних необластов сливаются. В конечном итоге на поверхности тела формируется синцитиальный слой, лишенный ресничек. Этот слой может формировать шипики, которые полностью находятся внутри цитоплазмы, их основания прикрепляются к базальной пластинке, а вершины торчат наружу, выпячивая апикальную мембрану тегумента. Как и покровы других животных, тегумент (неодермис) выполняет защитную и опорную функции. В числе других функций тегумента следует назвать экскрецию, осморегуляцию и функцию поглощения веществ (за счет пиноцитоза). При интенсификации этой последней функции тегумент образует разнообразные выросты - микроворсинки. Специфическими для паразитов являются функции защиты от воздействия организма хозяина - иммунного воздействия и пищеварительных ферментов. Для мышечной системы характерно - в сравнении с турбелляриями - отсутствие диагональных мышц и в целом более слабое развитие. Пищеварительная система начинается ротовым отверстием, которое расположено в ротовой присоске, Далее следует сосательная глотка, устроенная по типу бульбовидной глотки прямокишечных турбеллярий. За глоткой находится короткий пищевод, развевляюящийся на две слепо замкнутые ветви кишечника. В типовом случае эти две ветви доходят почти до заднего конца тела червя и более не ветвятся. У мелких видов ветви могут быть более короткими, доходя примерно до середины тела. Для крупных видов характерно, что две названные ветви в свою очередь сильно ветвятся. Это решает проблему транспортировки питательных веществ. Отметим, что у крупных видов ветвистую форму могут принимать гонады, а желточники представлены мелкими гроздевидными фолликулами практически у всех трематод. Это разветвеление органов увеличивает их поверхность, что опять-таки решает проблемы получения питательных веществ. Пищей трематодам служат разные субстанции тела хозяина: полупереваренное или полностью переваренное содержимое кишечника, клеточный детрит, тканевая жидкость, кровь. Переваривание пищи происходит в просвете кишечника. Все мариты трематод в той или иной степени поглощают растворенные питательные вещества поверхностью тегумента. У представителей семейства Strigeidae имеется особый бокаловидный орган, специализированный на внекишечном поглощении пищи. В редчайших случаях пищеварительная система отсутствует (род Anenterontrema). |