Конспект лекций по экологии для заочников. Лекция Краткая история и предмет экологии
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Лекция 6. Теоретические аспекты 1. Экологическая эффективность. 2. Элементы теории сложности в экологии. 3. Концепция поддерживающей емкости. 4. Метаболизм и размеры особей. 5. Трофическая структура и трофическая функция экосистемы. 6. Экологические пирамиды. 7. Концентрация токсичных соединений при прохождении по пищевым цепям. Экологическая эффективность. Значительный интерес для эколога представляют отношения величин энергетического потока в разных точках пищевой цепи. Выраженные в процентах, эти отношения часто называют экологической эффективностью. В табл. 6.1 приведены некоторые из этих отношений и даны определения, в основе которых лежат представления об энергетических потоках. По большей части эти отношения могут использоваться применительно и к составляющим популяциям, и к целым трофическим уровням. Таблица 6.1. Различные типы экологической эффективности
Уже обсуждалась эффективность переноса между трофическими уровнями; Отношение PG /L обычно равно 1-5%, PG /LA равно 2-10%, и эффективность переноса продукции между вторичными трофическими уровнями составляет обычно 10-20%. Поскольку у теплокровных животных (эндотермов), постоянно поддерживающих высокую температуру тела, доля ассимилированной энергии, которая должна идти на дыхание, как минимум в 10 раз больше, чем у холоднокровных (экзотермов), у первых должна быть ниже эффективность продукции Р/A. Поэтому в "беспозвоночной" пищевой цепи эффективность переноса энергии между трофическими уровнями должна быть выше, чем в цепи с участием млекопитающих. Как правило, эффективность роста тканей и эффективность ассимиляции у животных связаны обратной зависимостью. Для травоядных животных обычно характерна более высокая эффективность Р/А , но более низкая А/I, чем для хищников. Многих озадачивала очень низкая первичная эффективность, характерная для интактных природных систем, в сравнении с высокими КПД электромоторов и других двигателей. Это привело к мысли о необходимости серьезно рассмотреть возможности увеличения эффективности процессов, происходящих в природе. На самом деде, долгоживущие, крупномасштабные экосистемы нельзя приравнивать в этом отношении к недолговечным механическим системам. Во-первых, в живых системах много "горючего" затрачивается на "ремонт" и самоподдержание, а при расчете КПД двигателей не учитываются амортизация и расходы энергии на ремонт. Во-вторых, в определенных условиях быстрый рост может иметь большее значение для выживания, чем максимальная эффективность использования "горючего". (Простая аналогия: автомобилисту может быть важнее быстро достичь пункта назначения, развив высокую скорость, чем с максимальной эффективностью использовать бензин, придерживаясь малой скорости). Элементы теории сложности в экологии. Закон сложности. Важно понимать, что любое повышение эффективности биологической системы обернется увеличением затрат на ее поддержание. По мере того, как размеры и сложность системы увеличиваются, пропорционально, но еще быстрее увеличивается энергетическая стоимость поддержания структуры и функции системы. При удвоении размеров системы, как правило, более чем вдвое увеличивается количество энергии, которая должна отводиться на уменьшение энтропии, связанной с необходимостью сохранения структурной и функциональной сложности. При увеличении размера и сложности системы проявляется закон увеличения отдачи, или экономия, связанная с масштабами, например возрастает качество и устойчивость системы к нарушающим воздействиям. Однако при этом наблюдается также уменьшение отдачи или, расходы, связанные с масштабами, что объясняется увеличением стоимости откачивания неупорядоченности. Такое уменьшение отдачи внутренне присуще большим и сложным системам. Его можно сгладить, улучшив конструкцию системы, что увеличивает эффективность превращений энергии, но полностью устранить невозможно. Являются ли такого рода расходы, связанные с масштабами, неотъемлемым свойством экосистем, неизвестно, но по крайней мере часть возрастающей платы за сложность компенсируется преимуществами (экономия, связанная с масштабами). Метаболизм на единицу массы уменьшается с увеличением массы организма или биомассы леса, так что на единицу потока энергии удается поддерживать больше структурных образований. Добавочные функциональные цепи и петли обратной связи могут увеличить эффективность использования энергии и повторного использования веществ и могут повысить устойчивость или упругость по отношению к нарушающим воздействиям. Возможность проявления эмерджентных свойств, в том числе симбиоза между организмами, может увеличить общую эффективность. Но какие бы приспособления не возникали в системе, общая энтропия быстро увеличивается с ростом размеров. В результате все большая и большая доля общего потока энергии должна отклоняться на дыхание, связанное с поддержанием системы, в связи с чем все меньшая доля остается для нового роста. Когда расходы энергии на поддержание уравниваются с количеством доступной энергии, дальнейший рост прекращается, достигается теоретическая максимальная поддерживающая емкость. Концепцию поддерживающей емкости можно объяснить с помощью диаграмм типа показанных на рис.6.1. Рост размеров и сложности популяций, а также целых экосистем часто идет по S -образной или сигмоидной кривой. Обратим внимание на две точки на кривой роста: К - верхняя асимптота, соответствует максимальной поддерживающей емкости, т.е. количеству биомассы, которая может поддерживаться при данных условиях, а I - точка перегиба, где скорость роста максимальна. Уровень I называют максимальным постоянным выходом, либо оптимальной поддерживающей емкостью, способной сохраняться долгое время несмотря на изменчивость среды. Согласно теории сложности, собранная биомасса будет быстрее всего восстанавливаться на этом уровне. 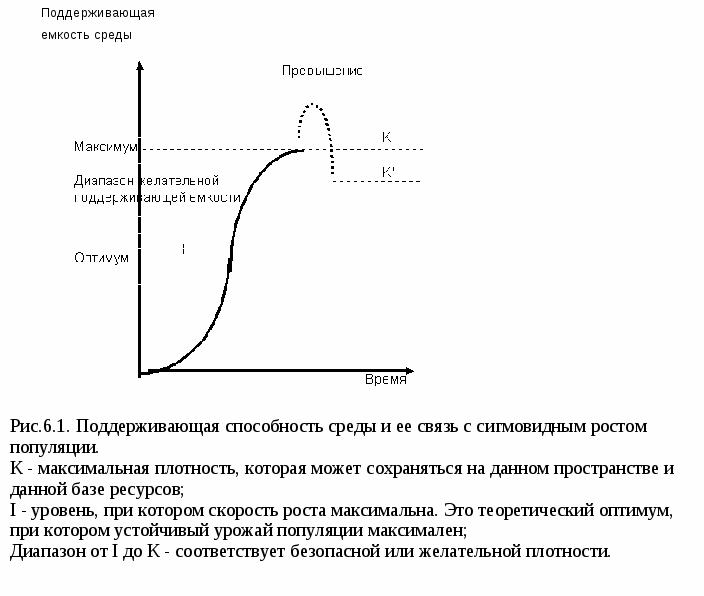 Сложность поддержания максимального уровня (К-уровня) в колеблющихся условиях реального мира состоит в том, что неизбежны отклонения от этого уровня: превышения вследствие инерции роста популяций и снижения, хотя бы временные, вследствие периодического уменьшения доступных ресурсов, например при засухе. Когда уровень становится выше максимального, и энтропия превосходит способность системы рассеивать ее, наблюдается уменьшение размеров или "крушение". Если при этом пострадала производительная способность среды, сам уровень К может временно снизиться (до уровня Кў). Безопасный допуск уровня максимальной поддерживающей емкости очень невелик. С точки зрения долгосрочной безопасности и стабильности уровень желательной поддерживающей емкости находится где-то между К и I. Это диапазон поддерживающей емкости. Мировая проблема снабжения человечества продуктами питания заключается в том, что потребность в них сейчас приближается к максимальной производительной способности Земли при существующих технологических, политических, экономических и распределительных ограничениях. Любое сильное стрессовое воздействие, например война, засуха или болезнь, которое сократит урожаи хотя бы на год, может означать серьезное недоедание или голод для миллионов людей. Трофическая структура и трофическая функция экосистемы. Метаболизм и размеры особей. Размер урожая биомассы на корню, выраженный в общей калорийности или в общей сухой массе всех организмов, присутствующих в данный момент времени, который может поддерживаться постоянным потоком энергии через пищевую цепь, в значительной степени зависит от размера особей. Чем меньше организм, тем выше его удельный метаболизм (т.е. метаболизм, обмен веществ в пересчете на 1 г или 1кал биомассы) и тем меньше биомасса, которая может поддерживаться на данном трофическом уровне. Так "урожай" бактерий, имеющихся в любой данный момент, будет гораздо ниже "урожая" рыбы или млекопитающих, хотя эти группы, возможно, используют одинаковое количество энергии. Различные теории, объясняющие эту тенденцию, особое внимание обращают на роль процессов диффузии; действительно у крупных организмов площадь поверхности, через которую может идти диффузия, на грамм массы меньше, чем у мелких. Однако общепринятого объяснения связи между размером и метаболизмом пока нет. Следует помнить, что с увеличением размеров снижается не общий метаболизм особи, а удельный. Взрослому человеку требуется пищи больше, чем маленькому ребенку, но на 1 кг массы взрослый потребляет меньше пищи. Трофическая структура и экологические пирамиды. В результате взаимодействия энергетических явлений в пищевых цепях (потерь энергии при каждом переносе) и такого фактора, как зависимость метаболизма от размеров особи, каждое сообщество приобретает определенную трофическую структуру, которая часто служит характеристикой экосистемы (озера, леса, пастбища и т.д.). Трофическую структуру можно измерить и выразить либо урожаем на корню на единицу площади, либо количеством энергии, фиксируемой на единицу площади за единицу времени на последовательных трофических уровнях. Трофическую структуру и трофическую функцию можно изобразить графически в виде экологических пирамид. Основанием этих пирамид служит первый трофический уровень - уровень продуцентов, а последующие уровни образуют этажи и вершину пирамиды. Экологические пирамиды можно отнести к трем основным типам: 1) пирамида численностей, отражающая численность отдельных организмов; 2) пирамида биомассы, характеризующая общую сухую массу, калорийность или другую меру общего количества живого вещества ; 3) пирамида энергии, показывающая величину потока энергии и (или) "продуктивность" на последовательных трофических уровнях. Пирамиды численности и массы могут быть обращенными, или частично обращенными, т.е. основание может быть меньше, чем один или несколько верхних этажей. Так бывает, когда средние размеры продуцентов меньше размеров консументов. Можно сформулировать некое "экологическое правило": данные по численности приводят к переоценке значения мелких организмов, а данные по биомассе - к переоценке роли крупных организмов. Следовательно, эти критерии непригодны для сравнения функциональной роли популяций, сильно различающихся по отношению интенсивности метаболизма к размеру особей, хотя, как правило, биомасса все же более надежный критерий, нежели численность. Из трех типов экологических пирамид пирамида энергии дает наиболее полное представление о функциональной организации сообществ. Число и масса организмов, которых может поддерживать какой- либо уровень в тех или иных условиях, зависит не от количества фиксированной энергии, имеющейся в данное время на предыдущем уровне, а от скорости продуцирования пищи. В противоположность пирамидам численностей и биомассы, отражающим статику системы, т.е. характеризующим количество организмов в данный момент, пирамида энергии отражает картину скоростей прохождения массы пищи через пищевую цепь. На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей, и если учтены все источники энергии, то пирамида всегда будет иметь "правильную форму", как это диктуется вторым законом термодинамики. Концепция потока энергии не только позволяет сравнивать экосистемы между собой, но и дает средство для оценки относительной роли популяций в их биотических сообществах. Многие исследователи считают, что трофическая структура является фундаментальным свойством каждого сообщества, ибо после острого нарушения его трофическая структура возвращается к равновесию независимо от видового состава и быстрее его, т.е. трофическая структура обладает способностью восстанавливаться. И только если экосистема постоянно находится под стрессовым воздействием, трофическая структура может измениться по мере приспособления биотических компонентов экосистемы к хроническим нарушениям. Концентрация токсичных соединений при прохождении по пищевым цепям Распределение энергии не единственный количественный параметр, на который влияют события, происходящие в пищевых цепях. Некоторые вещества по мере продвижения по цепи не рассеиваются, а наоборот, накапливаются. Это, так называемое, концентрирование в пищевой цепи нагляднее всего демонстрируют некото-рые устойчивые радионуклиды и пестициды. Так было обнаружено, что коэффициент накопления (соотношение количества вещества в тканях и в окружающей среде) радиоактивного фосфора равен 2 миллионам. Таким образом "безопасные выбросы" в окружающую среду могут стать крайне опасными для высших звеньев пищевой цепи, к которым относится и человек. Подобный пример демонстрирует и ДДТ. Чтобы сократить численность комаров на Лонг-Айленде, болота много лет опыляли ДДТ. Специалисты по борьбе с насекомыми благоразумно не применяли таких концентраций, которые были бы непосредственно летальны для рыбы и других животных, но они не учли экологических процессов и длительного сохранения токсичных остатков ДДТ. Ядовитые остатки, адсорбированные на детрите, концентрировались в тканях детритофагов и мелких рыб и далее - в хищниках высшего порядка - таких, как рыбоядные птицы. Коэффициент концентрации для рыбоядных птиц составляет около 500 000. Принцип биологического накопления надо учитывать при любых решениях, связанных с поступлением загрязняющих веществ в среду. Многие небиологические факторы, однако, могут уменьшать или увеличивать коэффициент концентрации. Так человек получает меньше ДДТ, чем рыбоядные птицы, и частично это объясняется тем, что при обработке и варке пищи часть этого вещества удаляется. Хищная рыба же находится в особенно опасном положении, потому, что может получать ДДТ не только через пищу, но и прямо из воды через жабры. Лекция 7. Биосфера как глобальная экосистема 1. Понятие о биосфере. 2. Живое вещество биосферы. 3. Геохимическая работа живого вещества. Понятие о биосфере. Представление о биосфере как общепланетарной оболочке, охватывающей толщу тропосферы, гидросферы, осадочных (и возможно гранитных) пород литосферы, в ходе всей геологической истории Земли; как глобальной единой системе Земли, где весь основной ход геохимических и энергетических превращений определяется жизнью, было разработано в трудах В.И.Вернадского. Вернадский впервые указал на активную преобразующую деятельность древних и современных организмов в изменении облика нашей планеты. Грандиозные масштабы этого процесса позволили ему развить учение о космической роли жизни в геологической истории Земли, что несомненно дает право считать его основателем учения о биосфере. Биосферой Вернадский назвал ту область нашей планеты, в которой существует или когда-либо существовала жизнь и которая постоянно подвергается или подвергалась воздействию живых организмов. Участие каждого отдельного организма в геологической истории Земли ничтожно мало. Однако живых существ на Земле бесконечно много, они обладают высоким потенциалом размножения, активно взаимодействуют со средой обитания и в конечном счете представляют в своей совокупности особый, глобальных масштабов фактор, преобразующий верхние оболочки Земли. Значение организмов обусловлено их большим разнообразием, повсеместным распространением, длительностью существования в истории Земли, избирательным характером биохимической деятельности и исключительно высокой химической активностью по сравнению с другими компонентами природы. Биосфера, таким образом, это та область Земли, которая охвачена влиянием живого вещества. С современных позиций биосферу рассматривают как наиболее крупную экосистему планеты, поддерживающую глобальный круговорот веществ. Современная жизнь распространена в верхней части земной коры (литосфере), в нижних слоях воздушной оболочки Земли (атмосфере) и в водной оболочке Земли (гидросфере). Для обозначения совокупности всего живого на Земле вместе с его непосредственным окружением и ресурсами введем термин "современная биосфера" или "экосфера". Экосфера непрерывной оболочкой одевает земной шар, а ее протяженность по вертикали меняется от долей метра - в областях чрезвычайно скудной жизни (арктические и антарктические пустыни) - до тысяч метров. Нижняя граница экосферы ограничена прежде всего температурой горных пород и подземных вод, которая постепенно возрастает с глубиной и на уровне 1,5 - 15 км уже превышает 100°С. Поэтому вглубь Земли живые организмы проникают на небольшое расстояние. Самая большая глубина, на которой в породах земной коры были обнаружены бактерии, составляет 4 км. В нефтяных месторождениях на глубине 2 - 2,5 км бактерии регистрируются в значительном количестве. В океане жизнь распространена до более значительных глубин и встречается даже на дне океанических впадин в 10 - 11 км от поверхности, так как температура там около 0°С. Однако по Вернадскому нижнюю границу биосферы следует проводить еще глубже. Постепенно накапливающиеся в океане гигантские толщи осадочных пород, происхождение которых связано с деятельностью живых существ - это тоже часть биосферы. В соответствии с динамическими процессами в земной коре осадочные породы постепенно вовлекаются в глубь ее, метаморфизируясь под действием высоких температуры и давления. Метаморфические породы земной коры, происходящие из осадочных, в конечном итоге также производные жизни. Верхняя граница жизни в атмосфере определяется нарастанием с высотой ультрафиолетовой радиации. На высоте 25 - 27 км большую часть ультрафиолетового излучения Солнца поглощает находящийся здесь тонкий слой озона - озоновый экран. Все живое, поднимающееся выше защитного слоя озона, погибает. Атмосфера же над поверхностью Земли насыщена многообразными живыми организмами. Споры бактерий и грибов обнаруживают до высоты 20 - 22 км, но основная часть аэропланктона сосредоточена в слое до 1-1,5 км. Хотя процессы жизнедеятельности современных организмов сосредоточены только в экосфере, влияние живого вещества (современного или существовавшего в прошлом) ощущается далеко за ее пределами. Именно поэтому биосфера Вернадского (как область существования всех былых экосфер) простирается далеко за пределы современной экосферы, охватывая по вертикали слой толщиной 40 - 50 км. Приблизительная масса биосферы составляет 0,05% массы Земли, а ее объем 0,4% объема планеты. Структура биосферы представляет собой сложную многокомпонентную систему - совокупность газообразной, жидкой, твердой и биологической организаций. Она характеризуется строгой организованностью, биологическим равновесием численности и взаимной адаптированностью составляющих ее организмов. Вернадский подчеркивал, что биосферу нужно рассматривать как целостную геологическую оболочку Земли, весьма сложную саморегулирующуюся систему, состоящую из живого вещества и неживой материи. Всю совокупность организмов на планете Вернадский назвал живым веществом. Косное вещество, по Вернадскому, это совокупность тех веществ в биосфере, в образовании которых живые организмы не участвуют - т.е. горные породы магматического, неорганического происхождения, видоизмененные живыми организмами вещества космического происхождения, космическая пыль, метеориты. Биогенное вещество создается и перерабатывается жизнью, совокупностями живых организмов. Это источник чрезвычайно мощной потенциальной энергии (каменный уголь, гумус почв, нефть, битумы, торф и т.п.). После образования биогенного вещества живые организмы в нем малодеятельны. Особой категорией является биокосное вещество. Вернадский определял, что оно "создается в биосфере одновременно живыми организмами и косными процессами, представляя системы динамического равновесия тех и других". Организмы в биокосном веществе играют ведущую роль. Биокосное вещество планеты - это почвы, кора выветривания, все природные воды, свойства которых зависят от деятельности на Земле живого вещества. |