Конспект лекций по экологии для заочников. Лекция Краткая история и предмет экологии
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Единицы измерения энергии. Хотя в системе СИ энергию измеряют в джоулях, все еще широко применяется и традиционная единица - калория. Определения для обеих единиц даны в таблице 3.1. Таблица 3.1. Единицы энергии
Энергетические характеристики среды. Для описания "поведения" энергии в экосистемах подходит понятие "линейный поток энергии", поскольку в отличие от циклического движения веществ превращения энергии идут в одном направлении. Энергия, однажды использованная каким-либо организмом, превращается в тепло и утрачивается для экосистемы. Энергия может накапливаться, сберегаться (т.е. преобразовываться в более эффективные формы) и передаваться из одной части системы в другую, но она не может быть использована повторно. Живые замкнутые термодинамические системы невозможны. Каждый живой компонент, будь это организм или экосистема, должен получить от своей среды на входе постоянный приток энергии. Основной источник практически всей энергии на Земле - Солнце. На биосферу из космоса падает солнечный свет с энергией 2 кал·см-2·мин-1. Проходя через атмосферу, излучение экспоненциально ослабляется атмосферными газами и пылью и в ясный летний полдень до поверхности Земли может дойти не более 67% его энергии. Степень этого ослабления зависит от длины волны (частоты) света. Ультрафиолетовое излучение с длиной волны менее 300 нм, почти не проходит через озоновый слой, имеющийся в атмосфере на высоте около 25 км, и это очень удачно, поскольку такое излучение летально для незащищенной протоплазмы. Излучение в оптической области (видимый свет) ослабляется равномерно, а инфракрасное излучение поглощается в атмосфере неодинаково в зависимости от длины волны. Лучистая энергия, достигающая земной поверхности в ясный день, состоит примерно на 10% из ультрафиолетового излучения, на 45% - из видимого света и на 45% - из инфракрасного. Меньше всего при прохождении через плотные облака и воду ослабляется видимый свет. Следовательно, фотосинтез, нуждающийся именно в видимом свате, может идти и в пасмурные дни, и под слоем чистой воды некоторой толщины (10-20 м). Поступление солнечной энергии к автотрофному слою экосистемы за день обычно варьирует от 100 до 800 кал·см-2, в среднем составляя около 300-400 кал·см-2 (3000-4000 ккал·м-2) или 1,1-1,5 млн. ккал·м-2·год-1. Растительность сильно поглощает синие и красные лучи, а также дальнее инфракрасное излучение; зеленый свет поглощается не так сильно (мы видим в отраженном свете, поэтому растительность для нас преимущественно зеленых оттенков); ближнее инфракрасное излучение - очень слабо. Тенистая прохлада создается в лесу благодаря тому, что листва поглощает много видимого и дальнего инфракрасного излучения. Синий и красный свет (400-500 и 600-700 нм соответственно) поглощаются особенно сильно хлорофиллом, а энергия дальнего инфракрасного излучения - водой, содержащейся в листьях, и окружающими их водяными парами. Таким образом, зеленые растения эффективно поглощают синий и красный свет, наиболее важный для фотосинтеза. Как бы отбрасывая ближнее ИК- излучение, несущее основную часть солнечной тепловой энергии, листья наземных растений избегают опасного перегрева. Кроме того, листья охлаждаются за счет испарения, а водные растения, разумеется, охлаждаются водой. Судьба солнечной энергии, поступающей в биосферу, кратко рассмотрена в таблице 3.2. Таблица 3.2. Рассеяние энергии солнечного излучения ( в % от годового поступления в биосферу). (По Hulbert, 1971.)
Хотя всего лишь около 1% энергии переходит в пищу и другую биомассу, те примерно 70%, которые преобразуются в тепло, уходят на испарение, осадки, ветер и т.д., не теряются зря, т.к. эта энергия поддерживает температуру и приводит в действие системы погоды и круговорот воды, необходимые для жизни на Земле. Другой энергетический компонент среды обитания - тепловое излучение. Оно исходит от всех поверхностей и тел, температура которых выше абсолютного нуля. Это не только почва, вода и растения, но и облака, излучающие вниз, на экосистемы, значительные количества тепловой энергии. Потоки длинноволнового излучения, разумеется, распространяются беспрестанно и во всех направлениях, а солнечный компонент (прямая солнечная радиация) имеет четкую направленность и поступает только днем. Следовательно, количество тепловой энергии, получаемой со всех сторон за сутки летом животным на открытом пространстве или листом растения, может в несколько раз превышать направленное прямо вниз излучение Солнца. Кроме того, тепловая энергия поглощается биомассой полнее, чем солнечное излучение. Условия существования организмов (температура, скорость испарения воды, движения воздуха и воды) определяются общим потоком излучения, но для продуктивности экосистемы и для круговорота биогенных элементов в ней важнее суммарное прямое солнечное излучение, попадающее на автотрофный ярус экосистемы, т.е. солнечная энергия, получаемая зелеными растениями за недели, месяцы, весь год. Этот приток первичной энергии приводит в действие все биологические системы. Лекция 4. Энергия и продуктивность 1. Концепция продуктивности. 2. Энергетическая классификация систем. 3. Концепция градиента от субсидии до стресса. Концепция продуктивности. Первичная продуктивность экологической системы, сообщества или любой их части определяется как скорость, с которой солнечная энергия усваивается организмами-продуцентами (главным образом зелеными растениями) в процессе фотосинтеза или хемосинтеза, накапливаясь в форме органических веществ. В процессе производства органического вещества выделяют четыре последовательных уровня, или ступени: 1. Валовая первичная продуктивность - это общая скорость фотосинтеза, включая те органические вещества, которые за время измерений были израсходованы на дыхание ("валовый фотосинтез", "общая ассимиляция"). 2. Чистая первичная продуктивность - скорость накопления органического вещества в растительных тканях за вычетом того органического вещества, которое использовалось при дыхании растений за изучаемый период ("наблюдаемый фотосинтез", "чистая ассимиляция"). 3. Чистая продуктивность сообщества - скорость накопления органического вещества, не потребленного гетеротрофами (т.е. чистая первичная продукция минус потребление гетеротрофами) за учетный период. 4. Вторичная продуктивность - скорость накопления энергии на уровнях консументов. Высокие скорости продуцирования наблюдаются в естественных и искусственных экосистемах там, где физические факторы благоприятны, и особенно при поступлении дополнительной энергии извне, уменьшающем собственные затраты на поддержание жизнедеятельности. Такая дополнительная энергия может поступать в разной форме: в тропическом дождевом лесу - в форме работы ветра и дождя, в эстуарии - в форме энергии прилива, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животным. Оценивая продуктивность экосистемы, необходимо учитывать как утечки энергии, связанные со сбором урожая, загрязнением среды, неблагоприятными климатическими условиями и другими типами стрессовых воздействий, способствующих отведению энергии от процесса продукции, так и поступления энергии, которые увеличивают продуктивность, компенсируя потери тепла при дыхании (при "откачивании неупорядоченности"), необходимые для поддержания биологической структуры. Всякий источник энергии, уменьшающий затраты на самоподдержание экосистемы и увеличивающий ту долю энергии, которая может перейти в продукцию, называется вспомогательным потоком энергии или энергетической субсидией. Энергетическая классификация систем. Источник и качество доступной энергии в той или иной степени определяют видовой состав и численность организмов, характер функциональных процессов, протекающих в экосистеме, и процессов ее развития, а также образ жизни человека. Энергия - общий знаменатель и исходная движущая сила всех экосистем, как сконструированных человеком, так и природных, следовательно, логично принять энергию за основу для первичной классификации экосистем. Удобно выделить на этой основе четыре фундаментальных типа экосистем. 1. Природные, движимые Солнцем, несубсидируемые; 2. Природные, движимые Солнцем, субсидируемые другими естественными источниками энергии; 3. Движимые Солнцем и субсидируемые человеком; 4. Индустриально-городские, движимые топливом (ископаемым, другим органическим или ядерным). Энергетическая классификация основана на свойствах среды на входе, она коренным образом отличается от биомной классификации, основанной на внутренней структуре экосистем, но вместе с тем и дополняет ее. Природные системы, в основном или полностью зависящие от прямого солнечного излучения, можно назвать движимыми Солнцем несубсидируемыми экосистемами. Они совсем или почти не получают дополнительной энергии, помимо солнечного света. К числу таких экосистем можно отнести открытие океаны, крупные участки горных лесов, грасленды и большие глубокие озера. Часто на них накладываются и другие ограничения, например, нехватка элементов питания и воды. Поэтому хотя экосистемы этой обширной группы весьма различны, все они получают мало энергии (от 1000 - 10000 ккал·м-2·год-1) и имеют низкую продуктивность или способность выполнять работу. Организмы, живущие в таких системах, выработали замечательные адаптации к существованию на скудном пайке энергии и других ресурсов и к эффективному их использованию. Хотя мощность природных экосистем, относящихся к первой категории, не очень впечатляет, и они не способны поддерживать высокую плотность населения, тем не менее, такие экосистемы крайне важны, так как занимают огромные площади (одни лишь океаны - до 70 % площади земного шара). Весь комплекс движимых Солнцем природных экосистем крайне важен для человека, это по сути дела основной гомеостат, стабилизирующий и поддерживающий условия жизни на планете; именно здесь ежедневно очищаются большие объемы воздуха, возвращается в оборот вода, формируются климатические условия, измеряются крайности погоды и выполняется множество других полезных функций. Если помимо солнечного света могут быть использованы какие-то дополнительные источники энергии, плотность мощности может быть значительно повышена, порой даже на порядок величины (10000 - 40000 ккал·м-2·год-1). В этом случае несолнечная энергия частично заменяет солнечную, сокращая расходы на самоподдержание системы, и высвобождает солнечную энергию на производство органических веществ. Источники дополнительной энергии могут быть как естественными, так и искусственными. Для простоты классификации выделены категории: движимые Солнцем экосистемы с естественными и с искусственными энергетическими субсидиями. Вспомогательная энергия, увеличивающая продуктивность, может поступать в самых разнообразных формах, например, в тропическом дождевом лесу - в форме ветра и дождя, в небольшом озере - в форме потока воды из ручья, или поступающих с площади водосбора органических веществ и минеральных элементов. Прибрежная часть эстуария - хороший пример природной экосистемы с дополнительной энергией приливов, прибоя и течений. Поскольку приливы и течения воды способствуют более быстрому круговороту минеральных элементов питания и перемещению пищи и отходов, организмы в эстуарии могут, так сказать, сконцентрировать свои усилия на более эффективном превращении энергии Солнца в органическое вещество. Человек также давно научился изменять природу и использовать вспомогательные источники энергии для получения прямой выгоды, а его умение не только увеличивать продуктивность, но и направлять эту продуктивность на производство пищевых и волокнистых материалов, легко собираемых, перерабатываемых и используемых, постоянно растет. Наземные и водные агроэкосистемы - основные примеры систем движимых Солнцем и субсидируемых человеком. Высокая продуктивность поддерживается большими поступлениями энергии топлива (а при более примитивных системах сельского хозяйства - мышечных усилий человека и животных). Эта энергия тратится на возделывание, орошение и удобрение, селекцию и борьбу с вредителями. Самое продуктивное сельское хозяйство находится примерно на уровне самых продуктивных природных экосистем: по-видимому, верхний предел для любой постоянной, длительно функционирующей системы, основанной на фотосинтезе, составляет примерно 50000 ккал·м-2·год-1. Действительное различие между природными и искусственными экосистемами состоит лишь в распределении этого потока энергии. Человек старается направить как можно больше энергии на производство продуктов питания, которые он может немедленно использовать, стараясь уменьшить расход продукции на автотрофном и гетеротрофном уровнях и увеличить чистую первичную продукцию и чистую продукцию сообщества; а естественные сообщества, получающие дополнительную энергию от природных ее источников, дают наибольшую валовую продуктивность и обычно распределяют продукты фотосинтеза между многими видами и веществами, это, так называемая, стратегия повышения разнообразия в целях выживания. В экосистеме, движимой топливом высококонцентрированная потенциальная энергия топлива не просто дополняет, а заменяет солнечную энергию. При современных методах ведения городского хозяйства солнечная энергия в самом городе не только не используется, но становится дорогостоящей помехой, так как она нагревает бетон и способствует образованию смога. Важное свойство экосистем, движимых топливом - огромная потребность в энергии плотно населенных индустриально-городских районов, она по меньшей мере на 2-3 порядка больше того потока энергии, который поддерживает жизнь в естественных условиях. Килокалории энергии, ежегодно протекающие через квадратный метр индустриального города, исчисляется уже не тысячами, а миллионами (100000 - 3000000 ккал·м-2·год-1). Вот почему небольшая территория города может поддерживать высокую плотность населения. Рассматривая общую концепцию энергетических субсидий надо сделать еще одно замечание, фактор, который при одних условиях среды или при одном уровне поступлений увеличивает продуктивность, при других условиях среды или другом уровне поступлений может способствовать утечкам энергии, уменьшая продуктивность. Слишком много хорошего также вредит системе, как и слишком мало. Например, некоторые виды загрязнений - обработанные сточные воды, могут в зависимости от объема и периодичности сброса оказаться либо благоприятным фактором, либо источником стресса. Если обработанные сточные воды попадают в экосистему с постоянной умеренной скоростью, то они, доставляя в систему микроэлементы, могут способствовать повышению продуктивности, однако массовый их сброс через нерегулярные промежутки времени может почти полностью уничтожить систему как биологическую единицу. Концепция градиента от субсидии до стресса. Концепция градиента от субсидии до стресса иллюстрируется рис.4.1. Если нарушение на входе вредно, то реакция экосистемы должна быть отрицательной. Но если на вход поступают полезные вещества или энергия, то продуктивность (или другие меры эффективности) может увеличиться: такая экосистема и будет называться субсидируемой экосистемой. При увеличении поступления субсидий способность системы их усваивать может достичь предела; эффективность после этого снизится. Хорошая иллюстрация соотношений между величиной субсидий и стрессовым воздействием - реакция системы на повышенное внесение азотных удобрений.  Рис.4.1. Иллюстрация концепции градиента от субсидии до стресса. Лекция 5. Пищевые цепи, пищевые сети и трофические уровни 1. Принципы организации и типы пищевых цепей. 2. Компоненты "универсальной" модели потоков энергии. Принципы организации и типы пищевых цепей. Обратимся теперь к изучению той части общего потока энергии, которая проходит через биотические компоненты экосистемы. Перенос энергии пищи от ее источника - автотрофов (растений) через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой цепью. Пищевые цепи можно разделить на два типа: пастбищная цепь, которая начинается с зеленого растения и идет далее к пасущимся растительноядным животным (т.е. к организмам, поедающим живые растительные клетки или ткани) и к хищникам (организмам, поедающим животных; и детритная цепь, которая от мертвого органического вещества идет к микроорганизмам, а затем к детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом, образуя так называемые пищевые сети. В сложных природных сообществах организмы, получающие свою энергию от Солнца через одинаковое число ступеней (звеньев пищевой цепи), считаются принадлежащими к одному трофическому уровню. Первый трофический уровень представлен первичными продуцентами или автотрофами; к ним относятся зеленые растения, которые способны использовать солнечный свет для образования химических соединений, богатых энергией. Первичные продуценты - это весьма важная часть сообщества, потому что практически все остальные организмы, входящие в его состав, прямо или косвенно зависят от снабжения энергией, запасенной растениями. Помимо первичных продуцентов в состав сообщества входят гетеротрофы, которые представлены консументами и деструкторами. Второй трофический уровень образуют растительноядные животные, называемые первичными консументами. Плотоядных, которые питаются растительно(траво-)ядными называют вторичными консументами или первичными хищниками; они занимают третий трофический уровень. Хищники, питающиеся первичными хищниками, в свою очередь, образуют четвертый трофический уровень и называются третичными консументами или вторичными хищниками. Точно также животные, потребляющие вторичных хищников, называются четвертичными консументами или третичными хищниками, они находятся на пятом трофическом уровне и т.д. Эта трофическая классификация относится к функциям, а не к видам как таковым. Поскольку многие животные всеядны и питаются как растениями, так и животными, т.е. одновременно получают энергию с нескольких разных трофических уровней, их невозможно отнести к определенному уровню. Принято считать, что такие организмы представляют сразу несколько трофических уровней, а их участие в каждом уровне пропорционально составу их диеты. В исследованиях структуры сообществ понятие трофического уровня оказалось чрезвычайно полезной абстракцией. Оно облегчает изучение потока вещества и энергии через сообщество и подчеркивает различия между взаимодействиями, которые протекают внутри трофических уровней и между ними. Принцип организации пищевых цепей и действия двух законов термодинамики можно уяснить, рассмотрев схему переноса энергии на рис.5.1. На этой схеме четырехугольники изображают трофические уровни, "трубы" - потоки энергии от каждого уровня или к нему. Как требует первый закон термодинамики, приток энергии уравновешивается ее оттоком, и каждый перенос энергии сопровождается ее рассеянием в форме недоступной для использования тепловой энергии (при дыхании), как того требует второй закон. 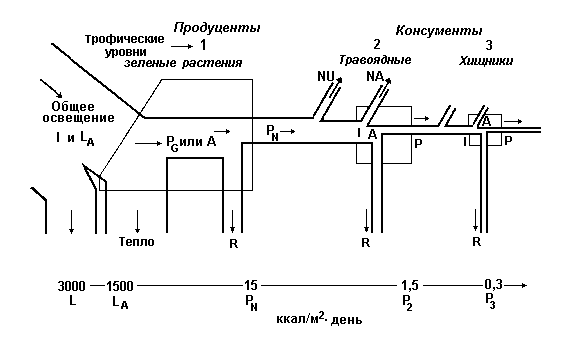 Рис.5.1. Упрощенная схема потока энергии, показывающая три трофическиих уровня (I, II и III) в линейной пищевой цепи (E.Odum, 1963.) I - общее поступление энергии; LA - свет, поглощаемый растительным покровом; PG - валовая первичная продуктивность; А - общая ассимиляция; PN - чистая первичная продукция; P2-3 - вторичная продукция (консументов); NU- неиспользуемая (накапливаемая или экспортируемая энергия); NA- неассимилированная консументами (выделенная с экскрементами) энергия; R - дыхание. Представленная схема потоков энергии на трех трофических уровнях сильно упрощена. Но она позволяет ввести принятые в литературе обозначения разных потоков и ясно показывает, что на каждом последующем уровне поток энергии сильно уменьшается независимо от того, рассматривается ли общий поток (I - общий поток энергии и А - общая ассимиляция) или компоненты P (продуктивность биомассы)и R (дыхание). Показано, что на первом трофическом уровне поглощается около 50% падающего света, а превращается в энергию пищи всего 1% поглощенной энергии, а также "двойной метаболизм" продуцентов (т.е. валовая и чистая продукция). Вторичная продуктивность (Р2, P3) на каждом последующем трофическом уровне консументов составляет около 10% предыдущей , хотя на уровне хищников эффективность может быть выше, скажем 20%. Если питательная ценность источника энергии велика (например, продукт фотосинтеза, извлекаемый или выделяемой прямо из растительных тканей), то эффективность переноса энергии может быть гораздо выше. Но поскольку и растения, и животные производят много трудноперевариваемого вещества (целлюлоза, лигнин, хитин), а также химические ингибиторы, препятствующие поеданию различными консументами, средняя эффективность переноса энергии между трофическими уровнями в целом составляет 20% и менее. Обратимся теперь к основным компонентам модели потока энергии. На рисунке 5.2 представлена модель, которую можно назвать "универсальной", поскольку она приложима к любому живому компоненту системы, будь то растение, животное, микроорганизм, особь, популяция или трофическая группа. Соединенные между собой, такие графические модели могут отразить, как уже показано (рис. 5.1) пищевые цепи или биоэнергетику экосистемы в целом. 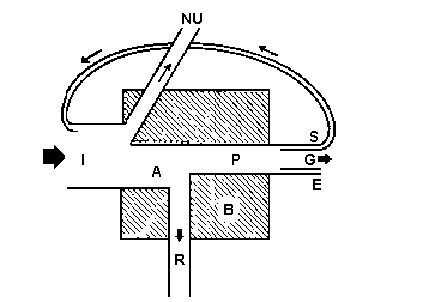 Рис.5.2. Компоненты "универсальной" модели экологического потока энергии. (E.Odum, 1968.) На рис.5.2 квадрат, обозначенный буквой B, изображает живую структуру, или биомассу, основного компонента модели. Желательно выражать биомассу в калориях, чтобы выявить связь между размерами потока энергии и одномоментной или средней биомассой. Общее поступление энергии - источник, обозначено буквой I. Для облигатных автотрофов - это свет, для облигатных гетеротрофов - органическая пища. Некоторые водоросли и бактерии могут использовать оба источника энергии, а многие из них нуждаются в определенном соотношении этих источников. Сходная ситуация наблюдается у лишайников и тех беспозвоночных животных, в которых содержатся водоросли-мутуалисты. В таких случаях канал притока на схеме потоков энергии можно разделить пропорционально использованию различных источников энергии либо можно разделить биомассу на отдельные резервуары. Тогда каждый резервуар (квадрат) будет соответствовать одному энергетическому (трофическому) уровню. Не вся энергия, поступающая в биомассу, подвергается превращению. Часть ее может просто пройти через биологическую структуру; например, пища может пройти через пищеварительный тракт, не включаясь в метаболизм, а часть энергии света проходит через растительность, не усваиваясь. Эта часть энергии обозначается через NU. Используемая, или ассимилируемая, часть на схеме обозначена буквой А. Отношение А и I, т.е. эффективность ассимиляции, широко варьирует. Оно может быть очень мало, как в случае усвоения света растениями (< 1%) или асси-миляции пищи у животных-детритофагов, или же очень велико, как в случае животных или бактерий, питающихся высококалорийной пищей, например сахарами и аминокислотами, что указывает на важность такого параметра, как качество ресурса. У автотрофов ассимилированная энергия (А) - это валовая продукция или валовый фотосинтез. Аналогичный компонент (А) у гетеротрофов - это пища, продуцированная кем-то другим. У высших животных величину А часто называют "метаболизированной энергией". Ключевая особенность этой модели - разделение ассимилированной энергии на компоненты P и R. Та часть фиксированной энергии А, которая окисляется и теряется в форме тепла, называется дыханием (R), а та часть, которая превращается в новое или принадлежащее другому виду органическое вещество, называется продукцией (P). У растений это чистая продукция, у животных - вторичная продукция. Компонент P - это энергия, доступная следующему трофическому уровню, в противоположность компоненту NU , который доступен и на данном трофическом уровне. Отношения R/P и R/B широко варьируют. Они имеют важное экологическое значение. В экосистеме отношение общего дыхания сообщества к его суммарной биомассе (R/B) можно рассматривать как отношение затрат энергии на поддержание жизнедеятельности к энергии, заключенной в структуре, или как меру термодинамической упорядоченности. Это "соотношение Шредингера" служит мерой экологического оборота. Если выразить R и B в калориях и отнести их к абсолютной температуре, то отношение R/B становится отношением прироста энтропии, связанного с поддержанием структуры, к энтропии упорядоченной части. В целом часть энергии, идущая на дыхание, или на поддержание структуры, велика в популяциях крупных организмов (люди, деревья) и в сообществах с большой биомассой на корню. При стрессовых воздействиях на систему R возрастает. Величина P, напротив, сравнительно велика в активных популяциях мелких организмов, например бактерий или водорослей, в молодых, быстро растущих сообществах и в системах, получающих энергетические дотации. Продукция может принимать различные формы. Три ее типа указаны на рисунке 5.2 (G - рост, или увеличение, биомассы, E - ассимилированное органическое вещество, выделяемое с экскрементами, секретируемое или извлекаемое, S- запас (например, жировые накопления, которые могут быть использованы позже). Обратный поток S можно назвать и "рабочей петлей", поскольку это та часть продукции, которая используется для обеспечения энергией в будущем (например, хищник использует энергию запасенных веществ, чтобы найти новую жертву, энергия экскрементов или выделений травоядного животного удобряет почву, поддерживая жизнь растений, которыми это животное питается). |