МЯДЕЛЕЦ ОБЩ.ГИСТ.. Литература для медицинских вузов все факультеты о. Д. Мяделец основы цитологии, эмбриологии и общей гистологии
 Скачать 4.75 Mb. Скачать 4.75 Mb.
|

|
Раздел гистологии, изучающий развитие, строение и функции тканей животного организма, называется общей гистологией. ОПРЕДЕЛЕНИЕ ПОНЯТИЯ «ТКАНЬ». Еще задолго до изобретения микроскопа анатомы обнаружили в организме человека и животных однородные части, в разных соотношениях входящие в состав органов и определяющие их строение. Первоначально эти части различали по чисто внешним признакам, выделяя мягкие, жидкие, волокнистые, клетчатые (построенные из камер) части. Термин «ткань» впервые применил английский ученый Н. Грю в 1671 г. в книге "Начала анатомии растений". При препарировании растений он обнаружил, что их структура напоминает структуру текстильной ткани. Иногда нечто похожее обнаруживалось и при препаровке животного организма. Поэтому в период оформления гистологии как самостоятельной науки понятие "ткань" стало использоваться как представление о простых системах организма. С этого времени начал формироваться новый уровень микроскопического изучения организма — тканевой. За более чем 200-летний период изучения тканей было предложено огромное количество определений понятия «ткань». Одно из первых научных определений было дано в 1852 году А. Келликером: «Ткань — это комплекс элементарных составных частей, объединенных в одно морфологическое и физиологическое целое». В понятие «части» он включал клетки, синцитии, симпласты. Удачное для своего времени определение ткани дал русский советский гистолог А.А. Заварзин (1938): «Ткань есть филогенетически обусловленная система гистологических элементов, объединенных общей функцией, структурой и часто — происхождением". В последнее время интенсивно изучается так называемый дифферон-ный принцип организации тканей. Поэтому существует ряд современных определений ткани, основанных на представлениях о дифферонах. Клеточный дифферон — это совокупность клеточных форм, составляющих ту или иную линию дифференцировки от стволовой до терминально дифференцированной клетки. Начальной клеткой клеточ-ного дифферона является стволовая клетка. Следующую стадию гисто i-гического ряда образуют полустволовые, или коммитированные, клеть и которые в отличие от стволовых клеток могут дифференцироваться тольк. . i каком-то одном направлении. Третьей и самой многочисленной часты-дифферона являются дифференцированные, функционально активны, клетки. Наконец, четвертым компонентом являются старые, функционал ьч • неактивные клетки и постклеточные структуры (см. ниже). В качестве при мера можно рассмотреть дифферон зпителиоцитов эпидермиса — кератин*. цитов. Он включает в себя такие клетки на последовательных стадиях рал..' тия, расположенных на разных уровнях эпидермального пласта: базальный кератиноцит (стволовая и полустволовая клетки) > шиповатый кератиноцит—» зернистый кератиноцит—» блестящий кератиноцит—) роговая чешуйка (корнеоцит, являющийся постк.и точной структурой). Современные определения ткани в большинстве своем учитываю; дифферонный принцип организации тканей. Одно из таких определен! ,; сделано А.А. Клишовым (1981): «Ткани представляют собой мозаичную морфофункциональную систему взаимодействующих клеточных дифферо нов, различающихся по генезу, направлению и уровню дифференцирован клеток»-. Различают монодифферонные (состоят из одного дифферона) и поли дифферонные ткани. К первым относятся, например, сердечная мышечная ткань (содержит один дифферон кардиомиоцитов), гладкая мышечная ткань (имеется только дифферон гладких миоцитов), а примером вторсл" вида тканей является рыхлая волокнистая неоформленная соединительная ткань (РВНСТ), которая содержит диффсроны фибробластов, макрофа гов, тканевых базофилов, нлазмоцитов, жировых клеток и др. В полидиф феронных тканях выделяют основной дифферон (в РВНСТ это дифферон фибробластов) и второстепенные диффероны. Ткани представляют собой не простую сумму клеток и неклеточны-структур, а тканевую систему, в которой составляющие элементы тесп< взаимосвязаны между собой. ТКАНЕВЫЕ ЭЛЕМЕНТЫ Каждая ткань состоит из составных частей, или элементов, которьп называются тканевыми элементами. По современным представлениям, су шествуют три основных вида тканевых элементов: клетки, межклеточное (промежуточное) вещество и симпласты. Некоторые авторы относят к числу тканевых элементов также синцитий (Афанасьев Ю.И. и соавторы 1989; Быков В.Л., 1998) и постклеточные структуры (Быков В.Л., 1998) На рис. 7.1 показаны различные виды тканевых элементов. Определение понятия «клетка» дано в лекции по цитологии. Это главный, основной тканевой элемент, за счет деятельности клетки образуются остальные виды тканевых элементов. 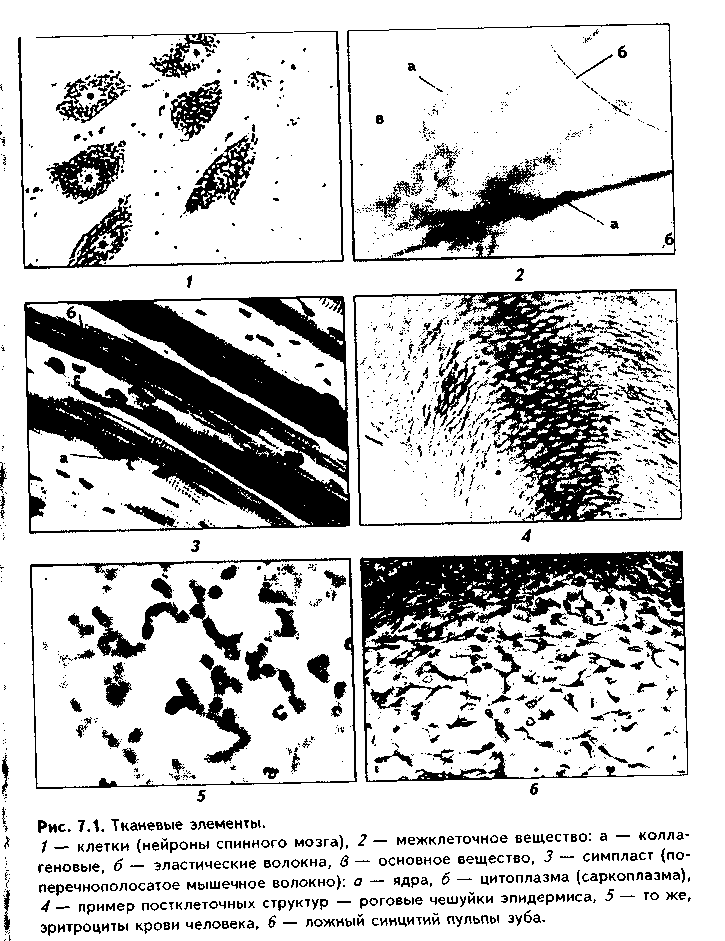 Межклеточное вещество — это тканевой элемент, который сип: зируется и секретируется особыми синтезирующими клетками и нахо/т , ■ -между клетками в составе ткани, составляя микросреду клеток. Mexi точное вещество состоит из основного (аморфного) вещества и воломm Основное вещество — это матрикс ткани, выполняющий метаболии кую, гомеостатическую, трофическую, регуляторную роль. Состой i n воды, белков, углеводов, липидов, минеральных веществ. Может бы п. состоянии золя (более жидкое) и геля (студнеобразное), а в костной i ■ ни — в минерализованном, твердом состоянии. Волокна выполни:' опорную, формообразующую функции, функцию эластичности, рсгу.п: руют функции клеток. Они делятся на коллагеновые, эластические, реш кулярные. Межклеточное вещество является тканевым элементом сосдп нительных тканей, и его строение более подробно будет изучено в com ветствующем разделе. Симпласт — это участок протоплазмы, ограниченный нлазмолсммон и содержащий большое количество ядер. Симпласты образуются путем слм яния клеток в отличие от многоядерных клеток, которые возникают в хо/н многократных делений клеток без цитотомии. Например, миосимплас! (поперечнополосатое мышечное волокно) образуется в эмбриогенезе путем слияния клеток миобластов. Второй пример симпластов — симпластотро фобласт хориона. В зарубежной литературе термин "симпласт" практичее ки не используется, вместо него применяются термины "многоядерн,: i клетка" или "синцитий". Синцитий. В отечественной гистологической литературе под синцитш' понимают совокупность клеток отросчатой формы, соединенных друг. другом цитоплазматическими мостиками. Различают "ложные" и "истинные" синцитии. В "ложных" синцитиях между отростками контактирую щих клеток имеются перерывы, представленные двумя клеточными пир < леммами и типичными контактами между ними. Примерами такого сим цития являются ретикулярная ткань, эпителий тимуса и пульпы эмалсво го органа развивающегося зуба. Единственным примером "истинного" сип цития являются развивающиеся мужские половые клетки. Синцитий п симпласт иногда называют надклеточными структурами. Постклеточные структуры. Это такие производные клеток, которые и результате терминальной дифференцировки утратили многие важнейшие признаки клеток: способность к репродукции, во многом обмен веществ и энергии и др. Данное обстоятельство связано с потерей клеточного ядра и резкой редукцией цитоплазматических органелл. Одновременно постклеточные структуры получили свойства, которые позволяют им в течение oi-раниченного времени выполнять некоторые узкоспецифические функции (функцию). К постклеточным структурам относятся эритроциты, тромбоциты, роговые чешуйки эпидермиса, волос, ногтей. ОБЩИЕ ФУНКЦИИ МНОГОКЛЕТОЧНЫХ КАК ОСНОВА ВОЗНИКНОВЕНИЯ ТКАНЕЙ В ФИЛО- И ОНТОГЕНЕЗЕ Каждый организм имеет некоторую сумму функций, которые обеспечи-Иают его существование. Прежде всего сюда относятся функции внутреннего и внешнего обмена. Несомненно, что эти функции должны были пер-Мыми появиться в филогенезе. У примитивных животных они играют основную роль. Если представить себе таких животных в виде плотного скопления клеточных элементов, то эти элементы окажутся в разных положениях по отношению к внешней среде. Лежащие на поверхности клетки непосредственно соприкасаются с внешней средой. Клетки, располагающиеся внутри, отделены от внешней среды и источников питания наружными клетками. Они получают питательные вещества через поверхностные клетки, т.е. оказываются в худших условиях питания. Однако вместе с тем они защищены от внешних воздействий поверхностными клетками. Па почве этих взаимоотношений и возникла, вероятно, внутренняя среда организма, отделенная от внешней среды барьером поверхностных клеточных элементов. Отсюда возникает вывод, что основными, а следовательно, и наиболее общими функциями всякого многоклеточного организма должны являться: 1. Барьерная, пограничная функция, т. е. функция внешнего обмена. 2. Функция внутреннего обмена, т. е. функция организации внутренней среды, опорно-трофическая функция. Поскольку всякая функция обеспечивается тем или иным морфологическим компонентом, эволюция этих функций должна была сопровождаться развитием двух основных типов тканей. Такими типами явились: 1. Пограничные, или барьерные, ткани. 2. Ткани внутренней среды. ПОГРАНИЧНЫЕ (БАРЬЕРНЫЕ), ИЛИ ЭПИТЕЛИАЛЬНЫЕ, ТКАНИ имеют плотное строение в форме сплошных клеточных пластов, причем клетки, составляющие эти пласты, полярно дифференцированы, т.к. одной своей поверхностью обращены к внешней, а другой — к внутренней среде. ТКАНИ ВНУТРЕННЕЙ СРЕДЫ, наоборот, состоят из клеток, не имеющих полярности. Кроме того, в их состав входит межклеточное вещество. В процессе эволюции для обеспечения общего обмена организма появляется необходимость и в других функциях. Так, для того, чтобы в меньшей степени зависеть от источников питания, возникла необходимость для передвижения организма в окружающем пространстве, движения внутренних органов. Это потребовало возникновения сократимых, или мышечных, тканей. Усложнение строения животного потребовало усовершенствования его реактивности, возбудимости, а также иптегрированности всех составных частей организма, что, соответственно, потребовало возникнове-ния нервной ткани. Эти два типа тканей, в отличие от двух первых тли-и (ткани общего назначения), называются специализированными tk;i нями. В филогенезе они появились позже, чем ткани общего назначение и возникли на их основе. Таким образом, фундаментальными функциями многоклеточна организмов, приведшими к возникновению основных типов тканей, >п ляются: 1. Функция внешнего обмена, или пограничная, барьерная функция. 2. Функция внутреннего обмена (защитная, опорно-трофическлл функция). 3. Функция сократимости, движения. 4. Функция возбудимости, реактивности, интегративная функция. КЛАССИФИКАЦИЯ ТКАНЕЙ Первые классификации тканей, основанные на микроскопическом изучении строения и развития, были предложены в середине XIX век-,: (А. Гассаль, А. Келликер, Ф. Лейдиг). Согласно этим классификациям различали 4 типа тканей: эпителиальные ткани; соединительные ткани < кровью; нервная ткань; мышечные ткани. Советский гистолог А.А. Заварзин положил в основу классификации тканей эволюционный принцип, основанный на фундаментальных функни ях многоклеточных организмов, возникающих в процессе их развития (cv выше). Он разделил все ткани на следующие типы: 1. Ткани общего назначения: 1.1. Пограничные ткани. 1.2. Ткани внутренней среды. 2. Специализированные ткани: 2.1. Ткани мышечной системы. 2.2. Ткани нервной системы. Другим советским гистологом, Н.Г. Хлопиным, была предложена генетическая классификация тканей, т. е. классификация, в основу которой положены источники развития тканей. Эта классификация выглядит так. 1. ЭПИТЕЛИЙ 1.1. Эпидермальный тип. 1.2. Энтеродермальный тип. 1.3. Целонефродермальный тип. 1.4. Эпендимоглиальный тип. 1.5. Ангиодермальный тип. 2. СОЕДИНИТЕЛЬНАЯ ТКАНЬ И КРОВЬ 2.1. Соединительная ткань и лейкоциты. 2.2. Эритроциты. 2.3. Хорда и хордальный хрящ. 2.4. Мезенхима. 3. МЫШЕЧНАЯ ТКАНЬ 3.1. Миокард. 3.2. Мезенхимальная гладкая мышечная ткань. 3.3. Соматическая миотомная мышечная ткань. 3.4. Мионейральная ткань. 3.5. Миоэнидермальиая ткань. 4. НЕРВНАЯ ТКАНЬ Нейроны, нейроглия. Классификация Н.Г. Хлопина вскрывает гистогенетические связи между функционально и структурно различающимися тканями. Наибольшее распространение получили гистогенетические классификации эпителиальных и мышечных тканей. РАЗВИТИЕ ТКАНЕЙ В ЭВОЛЮЦИИ В ходе эволюции происходило возникновение, развитие и усложнение строения различных тканей. Ход эволюции тканей наиболее полно объясняют следующие теории. Теория параллельных рядов. А.А. Заварзин разработал теорию эволюции тканей, которая называется теорией параллельных рядов тканевой эволюции, или теорией параллелизма. Суть этой теории заключается в том, что в ходе эволюции в разных ветвях филогенетического дерева самостоятельно, независимо, параллельно возникали одинаково построенные ткани, выполняющие сходные функции. Например, соединительная ткань ланцетника и млекопитающих выполняет одинаковые функции и поэтому имеет общие черты строения. Теория параллельных рядов хорошо раскрывает причины эволюции тканей, а также возможности их адаптации. Теория дивергентного развития тканей. Н.Г. Хлопин предложил собственную оригинальную теорию эволюции тканей, которая называется теорией дивергентного развития тканей. Согласно этой теории, ткани в эволюции и онтогенезе развиваются дивергентно, то есть возникают из уже существующих тканей путем расхождения признаков, что ведет ко все возрастающему разнообразию тканей. Эта теория показывает, как в ходе дивергенции из одного эмбрионального зачатка образуются ткани, постепенно приобретающие все более выраженные различия в строении и функциях. Например, развивающиеся из кожной эктодермы эпидермис и многослойный плоский эпителий имеют больше сходств, чем различий, тогда как имеющие общий с ними источник развития эпителий аденогипофиза, эмаль зуба и др. разительно от них отличаются. Единая концепция эволюционного развития тканей. Теории А. \ Заварзина и Н.Г. Хлопина органично дополняют друг друга. Поэтому сои< i ские гистологии А.А. Браун, В.П. Михайлов объединили их в единую теп рию эволюции тканей, которая утверждает, что сходные тканевые струкп ры в различных ветвях филогенетического дерева возникли параллельно и ходе дивергентного развития. В развитие учения о тканях большой вклад внесли такие ученые-гиск > логи, как А.А. Максимов (исследования крови, кроветворения, соединитель ной ткани; предложил унитарную теорию кроветворения, охарактеризова i стволовую кроветворную клетку). Советский гистолог Б.И. Лаврентьев и А.С. Догель изучали нервную ткань, ее реактивные свойства, они относя! ся к числу ученых-основоположников и разработчиков нейронной теории. Г.К. Хрущов посвятил свои исследования реактивным свойствам крови, ;i В.Г. Елисеев и его ученики (Ю.И. Афанасьев, Н.А. Юрина, А.Ф. Суханоп. А.И. Радостина, Е.Ф. Котовский и др.) — гистофизиологии и реактивным свойствам соединительной ткани и крови. Изучению скелетной мышечном ткани посвящены фундаментальные труды А.Н. Студицкого и А.А. Клишо-ва, сердечной мышечной ткани — П.П. Румянцева. Лауреат Нобелевской премии испанский гистолог С. Рамон-и-Кахал основное внимание уделил изучению нервной ткани, ее реактивных свойств. Он — один из основопо ложников нейронной теории. Большой вклад в изучение нервной ткани внес также итальянский гистолог, лауреат Нобелевской премии К. Гольджи. ПОНЯТИЕ О СТВОЛОВЫХ И ДИФФЕРЕНЦИРОВАННЫХ КЛЕТКАХ ТКАНЕЙ В составе тканей могут быть стволовые и дифференцированные клетки. Стволовые, или камбиальные, клетки представляют собой самоподдерживающуюся популяцию редко делящихся клеток, способных давать потомков, дифференцирующихся в различных направлениях под влиянием микроокружения (факторов дифференцировки). Стволовые клетки имеют следующие свойства: 1. Способны поддерживать постоянство численности своей популяции за счет двух процессов: редких митозов и дифференцировки в более зрелые клетки (после деления стволовой клетки одна остается стволовой, вторая — дифференцируется). Особо следует подчеркнуть, что стволовые клетки митотически делятся редко, большую часть своей жизни пребывают в состоянии покоя (G„ или в продленном G,-периоде (при этом их хроматин конденсируется) и (при необходимости) вновь могут вступать в митоти-ческий цикл, давая полустволовые, интенсивно делящиеся клетки. 2. Это клетки небольших размеров, которые имеют высокое ядерно-ци-топлазматическое отношение: в цитоплазме их содержится небольшое ко-Личество оргаиелл общего назначения; геном стволовых клеток находится в дерепгзессированном состоянии; 3. Для стволовых клеток характерен аутосинтетический тип обмена веществ: они синтезируют вещества только для собственных целей, для самоподдержания. 4. Стволовые клетки, как правило, устойчивы к повреждающим факторам. Это качество обеспечивается плотной упаковкой хроматина (преобладание гетерохроматина) в период митотического покоя. Кроме того, во многих тканевых системах стволовые клетки защищены местоположением (например, кроветворные стволовые клетки находятся в полостях костей; стволовые клетки эпидермиса лежат на дне эпидермальных гребешков, эпителия кишечника — в криптах, желудка — в железах, находящихся в соединительной ткани слизистой оболочки). Кроме того, стволовые клетки эпидермиса содержат большое количество гранул меланина, поглощающего вредные для клеток ультрафиолетовые лучи. 5. Способны к дифференцировке в различных направлениях. В процессе дифференцировки наблюдается следующая последовательность стадий: стволовая клетка —» полустволовая клетка -» унипотентная предшественница —> властная клетка (активно делящаяся) -»дифференцирующаяся клетка —»дифференцированная клетка. Дифференцированные (специализированные) клетки — это клетки, которые приобрели окончательные черты строения, необходимые для выполнения специфических функций. Они имеют следующие свойства: 1. Не способны делиться. 2. У них деблокирована (экспрессирована) только та часть генома, которая обеспечивает выполнение специфических функций. 3. Имеют низкое ядерно-цитоплазматическое отношение (сильно развита цитоплазма, в которой преобладают специфические для каждого вида клетки органеллы). 4. Свойственен гетеросинтетический тип обмена веществ (синтезируют и секретируют вещества для нужд организма). 5. Дифференцированные клетки имеют специфические, необходимые для выполнения специфических функций черты строения, и тинкториаль-ные свойства: базофилию цитоплазмы, полярность, развитие тех или иных органелл, характерную клеточную поверхность, определенное соотношение между гетеро- и эухроматином ядра и т.д. ЭМБРИОНАЛЬНЫЙ ГИСТОГЕНЕЗ. ИСТОЧНИКИ РАЗВИТИЯ ТКАНЕЙ Источником развития тканей являются эмбриональные зачатки. В свою очередь, эмбриональные зачатки развиваются из зародышевых листков в процессе их дифференцировки. В результате формируется осевой комплот зачатков (см. эмбриологию). Процесс образования тканей в эмбриогенезе и тканевых зачатков называется эмбриональным гистогенезом. Механизмы гистогенеза достаточно сложны и включают следующие компоненты (пол робнее см. в разделе «Эмбриология»): 1. Деление клеток. 2. Рост клеток. 3. Запрограммированная гибель клеток (апоптоз). 4. Адгезия клеток и межклеточные взаимодействия. 5. Детерминация. 6. Дифференцировка. 7. Эмбриональная индукция. 8. Миграция клеток. 9. Сегрегация клеток. ПОНЯТИЕ О КАМБИАЛЬНЫХ И НЕКАМБИАЛЬНЫХ (СТАЦИОНАРНЫХ) ТКАНЯХ, МЕХАНИЗМАХ ИХ ГИСТОГЕНЕЗА В результате вышеизложенных механизмов гистогенеза образуются камбиальные и некамбиальные ткани. Камбиальными называются ткани, в которых на протяжении всего по стнатального онтогенеза есть стволовые, или камбиальные, клетки. Гисто генез таких тканей в упрощенном виде происходит так: одна часть клеток после деления подвергается детерминации, дифференцировке, специализа ции, после чего выполняет специфические функции. Другая часть клеток остается в недифференцированном состоянии, выполняя роль стволовых клеток. При старении и гибели первой группы клеток камбиальные клетки начинают делиться, затем одна из образовавшихся клеток дифферен цируется, специализируется и выполняет функцию вместо погибшей клет ки, а вторая остается стволовой. К камбиальным тканям относятся соединительные, эпителиальны^ мышечные ткани (за исключением сердечной). В свою очередь, камбиаль ные ткани делятся на две группы: камбиальные обновляющиеся и камбиальные растущие. В камбиальных обновляющихся тканях пул стволовы:. клеток остается на протяжении всей жизни (РВНСТ, кровь, некоторьк эпителиальные ткани). В других тканях (поперечнополосатая мышечная ткань, эпителий печени) пул стволовых клеток постепенно снижается д< очень низкого уровня или совсем исчезает при достижении дефинитивно;-строения. Объем таких тканей постепенно увеличивается за счет внутри клеточной регенерации. Это камбиальные растущие ткани. Некамбиальные ткани — это ткани, в которых нет камбиальных клеток. В ходе эмбрионального гистогенеза клетки этих тканей делятся до достижения необходимого количества, а затем перестают делиться, детерминируются, дифференцируются, специализируются и начинают выполнять специфические функции. При этом стволовых клеток в тканях не остается, и их регенерация осуществляется только на внутриклеточном уровне. Подобное разделение тканей на группы в соответствии с регенераторными потенциями перекликается с приведенной в главе 4 классификацией клеточных популяций Леблона (статические, растущие и обновляющиеся клеточные популяции). СИСТЕМООБРАЗУЮЩИЕ ФАКТОРЫ ТКАНИ Как подчеркивалось, каждая ткань представляет собой систему, состоящую из взаимосвязанных тканевых элементов. Каждый тканевой элемент выполняет в ткани определенные функции, объем которых может существенно изменяться под действием внешних и внутренних факторов. Возникает полезный приспособительный результат, или цель действия, для данной функционирующей структуры. В последующем происходит своеобразный отбор признаков, отвечающих за определенную функцию. Это имеет большое значение не только для адаптации, но и для эволюции тканей. Факторы, влияющие на системогенез, в том числе и тканевой системо-генез (гистогенез), называются системообразующими факторами (СОФ). Системообразующие факторы ткани могут иметь значения от +100% до -100%. Если СОФ принимает положительные значения, то, по выражению И.И. Шмальгаузена, формируется движущий отбор, положительно изменяющий структуру ткани. При отрицательных значениях СОФ отбор носит стабилизирующий характер, закрепляя достигнутые положительные изменения. Нулевое значение СОФ означает дестабилизирующее влияние на ткань. Под действием СОФ происходит воспроизведение структурных частей тканей. В количественном отношении оно может быть избыточным или недостаточным. В первом случае СОФ будет равен +100%, в последнем случае — имеет нулевые значения, что ведет к деградации ткани. ТКАНЕВОЙ ГОМЕОСТАЗ. АДАПТАЦИЯ И РЕГЕНЕРАЦИЯ ТКАНЕЙ. РАЗЛИЧНЫЕ ТИПЫ РЕГЕНЕРАЦИИ ТКАНЕЙ. МЕТАПЛАЗИЯ Тканевой гомеостаз — это совокупность процессов поддержания постоянства структурно-функциональной организации ткани. Он реализуется следующими механизмами. 1. Поддержание диффереицировки клеток. Ткань — сложное сочетай,! взаимодействующих клеточных дифферонов. Находясь в ткани в один;и > вых условиях существования, они тем не менеее сохраняют свои различив свою специфическую дифференцировку благодаря клеточной памяти. 2. Поддержание необходимого нормального количества клеток (мехл низмы см. вьшш). 3. Поддержание необходимого объема внеклеточного матрикса (мел. клеточного вещества). Внеклеточный матрикс сам включается в процессы гомеостазирования ткани, поддерживает дифференцированное состояли клеток, регулирует клеточное обновление и препятствует апоптозу. 4. Обеспечение оптимального протекания обмена веществ и энергии 5. Поддержание нормального уровня физиологической регенерации как на клеточном, так и на внутриклеточном уровне. Регуляция тканевого гомеостаза. Осуществляется тканеспецифическм-ми и общими механизмами. Тканеспецифическая регуляция включает все механизмы регуляции численности клеточных популяций (внутриклеточные — ядерные, цитоплазматические, мембранные регуляторы митоза и апоптоза; межклеточные — кейлонная регуляция пролиферации, действие индукторов и т.д.). К тканеспецифической регуляции относится также ре гуляция функциональной активности клеток, в том числе и клеток-продуцентов межклеточного вещества. Эта регуляция осуществляется внутри тканевыми факторами — медиаторами, цитокинами и т.д. К общим механизмам регуляции тканевого гомеостаза относятся не рвная, эндокринная и иммунная регуляция. Строение ткани всегда отражает ее функции. Весь организм, а вместе с ним любой орган и любая ткань постоянно приспосабливаются к непрерыв но изменяющимся условиям функционирования. Поэтому в реальной жизни мы имеем дело не с тканевым гомеостазом, а с тканевым гомеокинезом. Адаптация — это приспособление ткани к изменяющимся условиям внешней и внутренней среды. Адаптация имеет место тогда, когда в ткани или других иерархических системах нет выраженного повреждения, а имеет место только усиление действия внешнего фактора. При возникновении повреждения мы имеем дело с компенсаторными (восстановительными) реакциями. В этих случаях для возмещения дефекта включаются сохранившиеся части тканей. В жизненных ситуациях, однако, чаще возникает комбинация адаптивных и компенсаторных реакций, поэтому говорят о компенсаторно-приспособительных реакциях в тканях. Комбинации компенсаторно-приспособительных реакций в тканях, которые наблюдаются в организме для поддержания тканевого гомеостаза, разнообразны, но принципы их реализации однотипны. 1. Непрерывное изменение режима функционирования ткани в соответствии с колебаниями силы и частоты действия возмущающего фактора. Как правило, осуществление компенсаторно-приспособительных реакций сопряжено с усилением функций, основанным на структурном обеспечении. Значительно реже имеет место приспособительное ослабление функций, например, снижение секреции потовых желез в условиях действия низких температур для уменьшения теплоотдачи или охранительное торможение в нервной ткани. Однако чаще ослабление функций ткани свидетельствует о переходе в состояние покоя. Такое почти универсальное значение усиления функций как внешнего элемента любой компенсаторно-приспособительной реакции объясняется тем, что в процессе эволюции организм в основном подвергался возбуждающим и разрушительным, а не успокаивающим воздействиям внешней среды, и нейтрализация их была возможна только путем интенсификации функций тканей и органов. 2. Усиление функции возможно за счет прироста функционирующих структур ткани: клеток, внеклеточного матрикса, симпластов и др. Прирост количества функционирующих структур ткани возможен: а) за счет включения в функции не задействованных в данный момент компонентов ткани. В покое действует принцип так называемого попеременного (асинхронного) функционирования одноименных структур. Например, в сердечной мышечной ткани в покое активно сокращается только определенная часть кардиомиоцитов, другие в это время "отдыхают", в них происходит внутриклеточная регенерация и т.д. При усилении физической нагрузки отмечается аварийное включение в работу большего числа или всех кардиомиоцитов. б) прирост количества клеток в ткани возможен либо за счет увеличения митотической активности клеток, либо за счет уменьшения их апоп-тотической гибели, либо путем сочетания этих процессов. Количество внеклеточных структур увеличивается путем их усиленного биосинтеза. В состоянии покоя биосинтез межклеточного вещества уравновешен его распадом. При адаптации деятельность клеток-продуцентов межклеточного вещества (фибробластов) активируется, а клеток, его разрушающих (фиб-рокластов, см. "Соединительную ткань") — подавляется. После снятия нагрузки происходит обратный процесс. 3. Важный принцип компенсаторно-приспособительных изменений — принцип экономизации, целесообразности: в организме никогда не содержатся лишние, избыточные структуры. Он всегда предпочитает строить их заново. Поэтому, если действие возмущающих факторов не подкрепляется, то объем ткани после адаптивно-компенсаторных перестроек постепенно возвращается к исходному. 4. В тканях, образованных стационарными клеточными популяциями, структурная сторона компенсаторно-приспособительных процессов реализуется за счет гипертрофии и гиперплазии органелл либо за счет сочетания этих процессов. Это приводит к гипертрофии клеточных элементов и уменьшению влияния на них возмущающего фактора. Может включать' также такой фактор, как синхронизация функционирования внутри клеточных структур, для которых также действует принцип асинхронно!" функционирования. 5. Очень важный механизм адаптации ткани — увеличение скоростей биологических реакций. Диапазон этих изменений очень велик: в экстре мальных условиях их интенсивность возрастает в десятки—сотни раз. 6. Какой бы срочной ни была компенсаторно-приспособительная реак ция, она никогда не осуществляется на чисто функциональной основе, i всегда имеет под собой соответствующую структурную базу (Д.С. Сарки сов). Регенерация — это способность клеток, тканей, органов восстанавлн вать погибшие или утраченные части. Регенерация направлена на сохра нение определенного уровня структурно-функциональной организации ткани. Различают физиологическую и репаративную регенерацию. Физиологическая регенерация протекает в условиях нормы. В органш ме постоянно происходит старение и смерть клеток, и при помощи физио логической регенерации ткани поддерживают свое постоянство, клеточный гомеостаз. В норме между гибелью и восстановлением тканевых элементом существует динамическое равновесие. По топографическому признаку физиологическая регенерация делится на несколько видов: 1. Мозаичная регенерация. В данном случае регенерация осуществля ется во многих мозаично расположенных участках ткани. В этих же участ ках происходит и гибель стареющих элементов, т.е топография восстанон ления и гибели элементов ткани совпадают. Примером являются РВНСТ мезотелий, эндотелий. 2. Зональная регенерация. При ней клетки ткани делятся в одной зон» ткани, а погибают — в другой, т.е. существует территориальное разобщение между процессами гибели и восстановления элементов ткани. Приме ром являются многослойные эпителии, эпителий коры надпочечника и др. 3. Дистантная регенерация. В этом случае восстановление тканевы> элементов (клеток) происходит в одних органах, а их физиологическая смерть в других органах (пример — кроветворные ткани: эритроциты обра зуются в красном костном мозге, а погибают в селезенке; лейкоциты, обра зовавшись в костном мозге, разрушаются в различных органах и тканях). Репаративная регенерация — это возникновение новых или гипертро фия оставшихся элементов ткани в ответ на повреждение. В основе физи ологической и репаративной регенерации лежат одни и те же механизмы которые реализуются как на внутриклеточном, так и на клеточном уровне Поэтому различают внутриклеточную и клеточную регенерацию. Внутриклеточная регенерация — это регенерация органелл клеток, увеличение их числа и размеров (гиперплазия, гипертрофия и их сочетание). Клеточная регенерация — это деление клеток и увеличение их числа, в результате чего происходит замещение погибших клеточных элементов ткани. Регенераторные способности ткани зависят от того, есть или нет в ней камбиальные клетки. Некамбиальные ткани регенерируют только на внутриклеточном уровне. Камбиальные ткани могут сочетать как клеточный, так и внутриклеточный уровень регенерации. Известный патолог Д.С. Саркисов в зависимости от форм регенерации тканей предложил следующую их классификацию (таблица). РАДИОЧУВСТВИТЕЛЬНОСТЬ И РАДИОРЕЗИСТЕНТНОСТЬ ТКАНЕЙ Радиочувствительность — это чувствительность тканей к действию ионизирующей радиации, тогда как радиорезистентность — устойчивость их к этому воздействию, т.е. эти понятия противоположны по значению. Общая реакция ткани на облучение определяется несколькими факторами: 1) количеством стволовых клеток; 2) величиной клеточной пролиферации; 3) скоростью утилизации зрелых клеток в здоровом и облученном организме. Согласно правилу Бергонье—Требондо, клетки ткани тем более чувствительны к облучению, чем больше у них способность к пролиферации и чем они менее дифференцированы. Таким образом, радиочувствительность тканей зависит от типа клеточных популяций, их составляющих. Камбиальные обновляющиеся ткани обладают наибольшей радиочувствительностью. Эпителиальные ткани в целом обладают высокой радиочувствительностью и низкой радиорезистентностью. Особенно чувствительны к облучению многослойные эпителии. В то же время, однослойный кубический эпителий почек из-за очень низкой митотической активности достаточно радиоустойчив.  РВНСТ обладает повышенной радиочувствительностью. После обл> ii< ния в ней наблюдается интенсивная гибель клеток, особенно камбиальны Вместе с тем, способность РВНСТ к участию в воспалении сохраняем i даже при интенсивных дозах облучения. Весьма чувствительны к облуи нию кроветворные ткани, тогда как плотная волокнистая соединителым i ткань связок и сухожилий имеет высокую радиорезистентность. Сердечная мышечная ткань относится к наиболее радиорезистентимм тканям, т.к не содержит камбиальных клеток. Даже очень большие до м облучения не приводят к заметным морфологическим изменениям мжж.ц' да. Скелетная мышечная ткань в дефинитивном состоянии содержи i очень мало камбиальных клеток и также является радиорезистентной. () i нако влияние радиации проявляется при регенерации и связано с пораж< нием входящей в состав мышц РВНСТ. Гладкая мышечная ткань относи i ся к камбиальным и поэтому радиочувствительна. Высокой устойчивостью обладает нервная ткань (нейроциты). Один ^ радиационное поражение последней проявляется при регенерации нервны\ волокон, что связано с поражением соединительной ткани и клеток глпп а также сосудистыми нарушениями. ИЗМЕНЧИВОСТЬ ТКАНЕЙ. Строение тканей закреплено в геноме составляющих ее клеток и, в значительной мере, постоянно на протяже нии всей жизни организма. Вместе с тем, каждая ткань подвергается опре деленным изменениям, пределы которых ограничены. Эти изменения мп гут быть двух видов: 1. Возрастные изменения (уменьшение количества клеток, снижешп способности к их размножению и регенерации; снижение и нарушение оо мена веществ, дистрофические изменения межклеточного вещества и др.) Очень часто возрастные изменения сопровождаются атрофией ткани снижением ее объема и функциональной активности. Атрофия ткани я и ляется следствием атрофии и уменьшения размеров клеток, уменьшения числа клеток, объема межклеточного вещества или наступает при сочета нии этих изменений. 2. Изменения тканей в процессе адаптации к неблагоприятным во:: действиям: увеличение митотической активности клеток, гиперплазия и гипертрофия клеток, усиление синтеза межклеточного вещества и в ре зультате увеличение общего объема ткани — гипертрофия ткани, которая может наступить или при реализации одного из указанных явлений, или при их сочетании. При длительном действии неблагоприятных факторов может наблю даться метаплазия ткани — превращение одной разновидности ткани г другую, родственную разновидность. Метаплазия возможна только в преде лах одного типа ткани, возникшего из одного зародышевого листка, а точ нее, в пределах одного тканевого зачатка. Она чаще встречается в эпителиж* И и соединительной ткани, реже — в других тканях. Метаплазия эпителия Чите всего проявляется в виде перехода многорядного реснитчатого эпителия в многослойный плоский эпителий (например, в дыхательных путях При хроническом бронхите у курильщиков или при недостатке витамина А). Метаплазия соединительной ткани с образованием хряща и кости может наблюдаться в рубцах, стенке аорты, строме мышц. Метаплазии всегда предшествует пролиферация клеток, которые затем дифференцируются в нетипичные для нормы клетки. Обратный процесс, т. е. возвращение ткани к нормальному строению, называется проплазией. Метаплазия часто является щюдраковым состоянием. |